SARS-CoV-2 and COVID-19: An Evolving Review of Diagnostics and Therapeutics
This manuscript
(permalink)
was automatically generated
from greenelab/covid19-review@910dd7b
on March 30, 2021.
Snapshots of individual sections are available as preprints [1,2,3].
This in progress manuscript is not intended for the general public.
This is a review paper that is authored by scientists for an audience of scientists to discuss research that is in progress.
If you are interested in guidelines on testing, therapies, or other issues related to your health, you should not use this document.
Instead, you should collect information from your local health department, the CDC’s guidance, or your own government.
Authors
Halie M. Rando 0000-0001-7688-1770 · rando2 · tamefoxtime
Department of Systems Pharmacology and Translational Therapeutics, University of Pennsylvania, Philadelphia, Pennsylvania, United States of America; Department of Biochemistry and Molecular Genetics, University of Colorado School of Medicine, Aurora, Colorado, United States of America; Center for Health AI, University of Colorado School of Medicine, Aurora, Colorado, United States of America
· Funded by the Gordon and Betty Moore Foundation (GBMF 4552)
Casey S. Greene 0000-0001-8713-9213 · cgreene · GreeneScientist
Department of Systems Pharmacology and Translational Therapeutics, University of Pennsylvania, Philadelphia, Pennsylvania, United States of America; Childhood Cancer Data Lab, Alex’s Lemonade Stand Foundation, Philadelphia, Pennsylvania, United States of America; Department of Biochemistry and Molecular Genetics, University of Colorado School of Medicine, Aurora, Colorado, United States of America; Center for Health AI, University of Colorado School of Medicine, Aurora, Colorado, United States of America
· Funded by the Gordon and Betty Moore Foundation (GBMF 4552); the National Human Genome Research Institute (R01 HG010067)
Michael P. Robson 0000-0002-4859-0033 · mprobson
Department of Computing Sciences, Villanova University, Villanova, Pennsylvania, United States of America
Simina M. Boca 0000-0002-1400-3398 · SiminaB
Innovation Center for Biomedical Informatics, Georgetown University Medical Center, Washington, District of Columbia, United States of America
Nils Wellhausen 0000-0001-8955-7582 · nilswellhausen
Department of Systems Pharmacology and Translational Therapeutics, University of Pennsylvania, Philadelphia, Pennsylvania, United States of America
Ronan Lordan 0000-0001-9668-3368 · RLordan · el_ronan
Institute for Translational Medicine and Therapeutics, Perelman School of Medicine, University of Pennsylvania, Philadelphia, PA 19104-5158, USA
Sandipan Ray 0000-0002-9960-5768 · rays1987
Department of Biotechnology, Indian Institute of Technology Hyderabad, Kandi, Sangareddy 502285, Telangana, India
Lucy D'Agostino McGowan 0000-0001-7297-9359 · LucyMcGowan · LucyStats
Department of Mathematics and Statistics, Wake Forest University, Winston-Salem, North Carolina, United States of America
Anthony Gitter 0000-0002-5324-9833 · agitter · anthonygitter
Department of Biostatistics and Medical Informatics, University of Wisconsin-Madison, Madison, Wisconsin, United States of America; Morgridge Institute for Research, Madison, Wisconsin, United States of America
· Funded by John W. and Jeanne M. Rowe Center for Research in Virology
Anna Ada Dattoli 0000-0003-1462-831X · aadattoli · aadattoli
Department of Systems Pharmacology & Translational Therapeutics, Perelman School of Medicine, University of Pennsylvania, Philadelphia, PA 19104, USA
John P. Barton 0000-0003-1467-421X · johnbarton · _jpbarton
Department of Physics and Astronomy, University of California-Riverside, Riverside, California, United States of America
Jeffrey M. Field 0000-0001-7161-7284 · Jeff-Field
Department of Pharmacology, Perelman School of Medicine, University of Pennsylvania, Philadelphia, PA 19104, USA
Adam L. MacLean 0000-0003-0689-7907 · alavendelm · adamlmaclean
Department of Quantitative and Computational Biology, University of Southern California, Los Angeles, California, United States of America
Alexandra J. Lee 0000-0002-0208-3730 · ajlee21
Department of Systems Pharmacology and Translational Therapeutics, University of Pennsylvania, Philadelphia, Pennsylvania, United States of America
· Funded by the Gordon and Betty Moore Foundation (GBMF 4552)
Immunology Institute of the Icahn School of Medicine · ismms-himc
Immunology Institute of the Icahn School of Medicine
Fengling Hu 0000-0003-1081-5038 · hufengling · hufengling
Department of Biostatistics, Epidemiology and Informatics, University of Pennsylvania, Philadelphia, Pennsylvania, United States of America
Nafisa M. Jadavji 0000-0002-3557-7307 · nafisajadavji · nafisajadavji
Biomedical Science, Midwestern University, Glendale, AZ, United States of America; Department of Neuroscience, Carleton University, Ottawa, Ontario, Canada
· Funded by the American Heart Association (20AIREA35050015)
Elizabeth Sell 0000-0002-9658-1107 · esell17
Perelman School of Medicine, University of Pennsylvania, Philadelphia, Pennsylvania, United States of America
Jinhui Wang 0000-0002-5796-8130 · jinhui2
Perelman School of Medicine, University of Pennsylvania, Philadelphia, Pennsylvania, United States of America
Diane N. Rafizadeh 0000-0002-2838-067X · dianerafi
Perelman School of Medicine, University of Pennsylvania, Philadelphia, Pennsylvania, United States of America; Department of Chemistry, University of Pennsylvania, Philadelphia, Pennsylvania, United States of America
· Funded by NIH Medical Scientist Training Program T32 GM07170
Ashwin N. Skelly 0000-0002-1565-3376 · anskelly
Perelman School of Medicine, University of Pennsylvania, Philadelphia, Pennsylvania, United States of America; Institute for Immunology, University of Pennsylvania Perelman School of Medicine, Philadelphia, United States of America
· Funded by NIH Medical Scientist Training Program T32 GM07170
Marouen Ben Guebila 0000-0001-5934-966X · marouenbg · marouenbg
Department of Biostatistics, Harvard School of Public Health, Boston, Massachusetts, United States of America
Likhitha Kolla 0000-0002-1169-906X · likhithakolla · lkolla2018
Perelman School of Medicine, University of Pennsylvania, Philadelphia, Pennsylvania, United States of America
· Funded by NIH Medical Scientist Training Program T32 GM07170
David Manheim 0000-0001-8599-8380 · davidmanheim · davidmanheim
1DaySooner, Delaware, United States of America; Risk and Health Communication Research Center, School of Public Health, University of Haifa, Haifa, Israel
Soumita Ghosh 0000-0002-2783-2750 · soumitagh
Institute of Translational Medicine and Therapeutics, Perelman School of Medicine, University of Pennsylvania, Philadelphia, Pennsylvania, United States of America
James Brian Byrd 0000-0002-0509-3520 · byrdjb · thebyrdlab
University of Michigan School of Medicine, Ann Arbor, Michigan, United States of America
· Funded by NIH K23HL128909; FastGrants
YoSon Park 0000-0002-0465-4744 · ypar · yoson
Department of Systems Pharmacology and Translational Therapeutics, University of Pennsylvania, Philadelphia, Pennsylvania, United States of America
· Funded by NHGRI R01 HG10067
Vikas Bansal 0000-0002-0944-7226 · bansalvi · VikasBansal1989
Biomedical Data Science and Machine Learning Group, German Center for Neurodegenerative Diseases, Tübingen 72076, Germany
Stephen Capone 0000-0001-7231-1535 · scapone01
St. George’s University School of Medicine, St. George’s, Grenada
John J. Dziak 0000-0003-0762-5495 · dziakj1
Edna Bennett Pierce Prevention Research Center, The Pennsylvania State University, University Park, PA, United States of America
Yuchen Sun · kevinsunofficial
Department of Computer Science, University of Virginia, Charlottesville, VA, United States of America
Yanjun Qi 0000-0002-5796-7453 · qiyanjun
Department of Computer Science, University of Virginia, Charlottesville, VA, United States of America
Lamonica Shinholster 0000-0001-6285-005X · LSH2126
Mercer University, Macon, GA, United States of America
· Funded by the Center for Global Genomics and Health Equity at the University of Pennsylvania
Temitayo Lukan · tlukan
University of Pennsylvania, Philadelphia, PA, United States of America
Dimitri Perrin 0000-0002-4007-5256 · SystemsResearch · dperrin
School of Computer Science, Queensland University of Technology, Brisbane, Australia; Centre for Data Science, Queensland University of Technology, Brisbane, Australia
Serghei Mangul 0000-0003-4770-3443 · smangul1 · serghei_mangul
Department of Clinical Pharmacy, School of Pharmacy, University of Southern California, Los Angeles, CA, United States of America
Shikta Das 0000-0002-8291-2788 · shiktadas · shikta_das
C4X Discovery, London, United Kingdom; Medical Research Council LHA, Institute of Cardiovascular Studies, University College London, London, United Kingdom
Tiago Lubiana 0000-0003-2473-2313 · lubianat · lubianat
Department of Clinical and Toxicological Analyses, School of Pharmaceutical Sciences, University of São Paulo, São Paulo, Brazil
Joel D Boerckel https://orcid.org/0000-0003-3126-3025 · jboerckel · jboerckel
Department of Orthopaedic Surgery, Perelman School of Medicine, University of Pennsylvania, Philadelphia, PA, United States of America - Department of Bioengineering, University of Pennsylvania, Philadelphia, PA, United States of America
COVID-19 Review Consortium:
Vikas Bansal, John P. Barton, Simina M. Boca, Joel D Boerckel, Christian Brueffer, James Brian Byrd, Stephen Capone, Shikta Das, Anna Ada Dattoli, John J. Dziak, Jeffrey M. Field, Soumita Ghosh, Anthony Gitter, Rishi Raj Goel, Casey S. Greene, Marouen Ben Guebila, Fengling Hu, Nafisa M. Jadavji, Sergey Knyazev, Likhitha Kolla, Alexandra J. Lee, Ronan Lordan, Tiago Lubiana, Temitayo Lukan, Adam L. MacLean, David Mai, Serghei Mangul, David Manheim, Lucy D'Agostino McGowan, YoSon Park, Dimitri Perrin, Yanjun Qi, Diane N. Rafizadeh, Bharath Ramsundar, Halie M. Rando, Sandipan Ray, Michael P. Robson, Elizabeth Sell, Lamonica Shinholster, Ashwin N. Skelly, Yuchen Sun, Gregory L Szeto, Ryan Velazquez, Jinhui Wang, Nils Wellhausen
Authors are ordered arbitrarily.
Abstract
Since late 2019, Coronavirus disease 2019 (COVID-19) has spread around the world, resulting in the declaration of a pandemic by the World Health Organization (WHO).
This infectious disease is caused by the newly identified Severe acute respiratory syndrome-related coronavirus 2 (SARS-CoV-2).
Research on the SARS-CoV-2 virus and the disease it causes is emerging rapidly through global scientific efforts.
Short-term mitigation of viral impacts will require public health interventions, and long-term mitigation will require new diagnostic and therapeutic technologies.
The urgency of the pandemic has led to a rapidly emerging scientific literature on SARS-CoV-2 and COVID-19.
This manuscript represents a collaborative effort to organize and consolidate this body of literature.
We present information about the virus in the context of what is known about related viruses, describe the pathogenesis of COVID-19, and synthesize studies emerging about the diagnosis and treatment of COVID-19 alongside literature about related illnesses.
We summarize this emerging literature with an eye towards discussing elements of the disease that will be fundamental to efforts to develop interventions.
Our review is a collaboratively-authored, evolving document into which we seek to incorporate the ever-expanding body of information on the topic.
This document provides a snapshot as of October, 2020.
We continue to accept new contributions and anticipate future snapshots until technologies to mitigate the pandemic are widely deployed.
On January 21, 2020, the World Health Organization (WHO) released its first report concerning what is now known as the Coronavirus Disease 2019 (COVID-19) [4].
This infectious disease came to international attention on December 31, 2019 following an announcement by national officials in China describing 44 cases of a respiratory infection of unknown cause.
The first known cases were located in Wuhan City within the Hubei province of China, but the disease spread rapidly throughout China and subsequently around the world.
At the time of the WHO’s first situation report [4], 282 confirmed cases had been identified.
Most of these cases were in China, but one to two exported cases had also been identified in each of several neighboring countries (Thailand, Japan, and the Republic of Korea).
One week later, 4,593 confirmed cases had been identified, spanning not only Asia, but also Australia, North America, and Europe [5].
On March 11, 2020, the WHO formally classified the situation as a pandemic [6].
On April 4, 2020, the WHO reported that the global number of confirmed cases had surpassed one million [7].
As international attention remains focused on the ongoing public health crisis, the scientific community has responded by mobilizing resources and turning much of its attention to the virus and disease.
This rapid influx of information is disseminated by traditional publishing mechanisms, preprint servers, and press releases, which provide a venue for scientists to release findings without undergoing the formal publication process.
While having information available is valuable to efforts to understand and combat COVID-19, many contributions come from researchers across a wide range of fields who have varying degrees of experience working on coronaviruses and related topics.
The volume of information available, much of which has not gone through rigorous peer review, presents a significant challenge to individual efforts to keep abreast of the state of COVID-19 research [8].
However, research on these topics is proceeding so rapidly that any static review is likely to quickly become dated.
Our goal as a community is to consolidate information about the virus in the context of related viruses and to synthesize rapidly emerging literature centered on the diagnosis and treatment of COVID-19.
We used an open publishing framework, Manubot [9], to manage hundreds of contributions from the community to create a living, scholarly document.
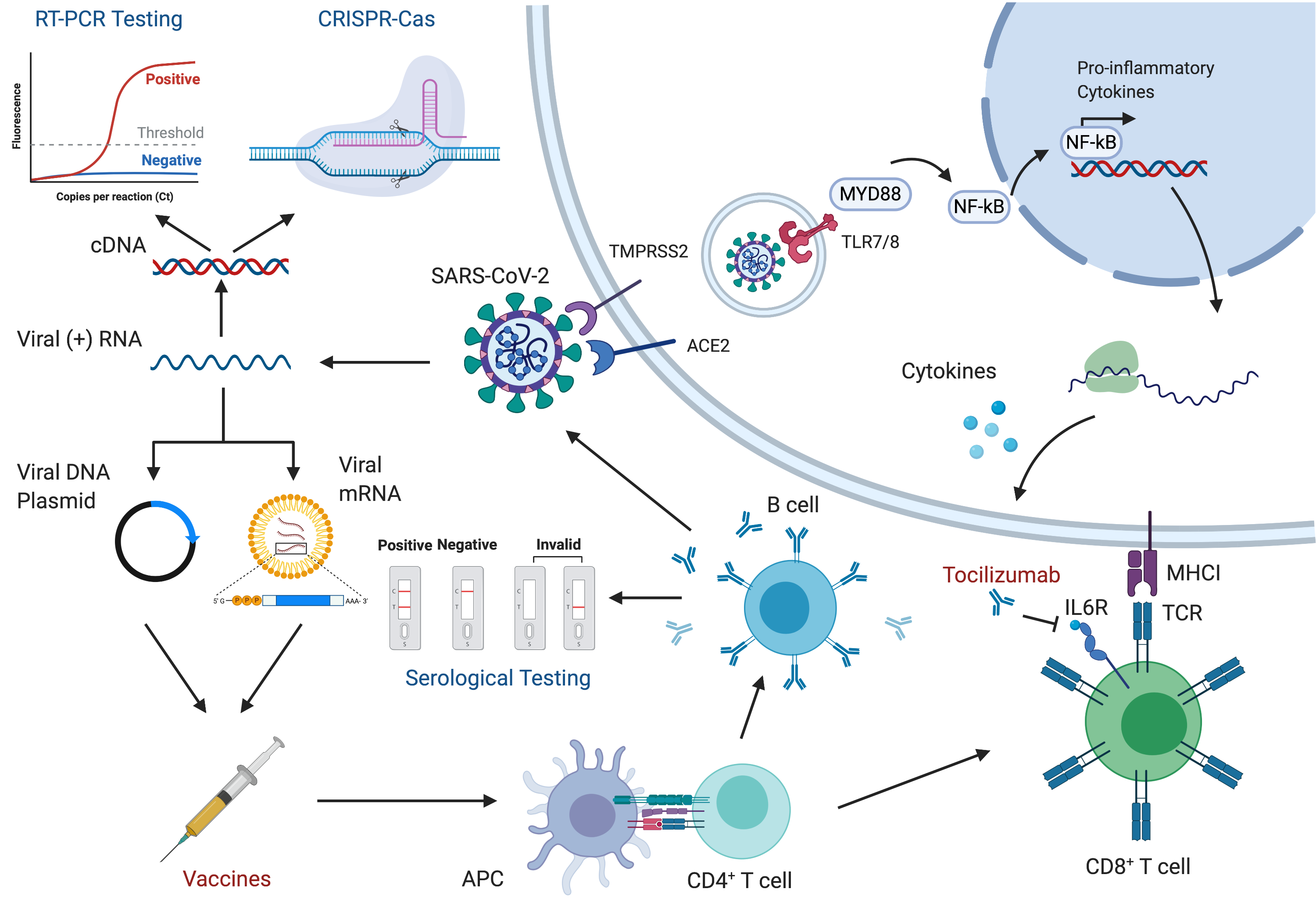
We designed software to generate figures, such as , that automatically update using external data sources.
Our primary goal is to sort and distill informative content out of the overwhelming flood of information [8] and help the broader scientific community become more conversant on this critical subject.
Thus, our approach has been to develop a real-time, collaborative effort that welcomes submissions from scientists worldwide into this ongoing effort.
This document represents the first snapshot, which aims to reflect the state of the field as of October, 2020.
We plan to refine and expand this document until technologies to mitigate the pandemic are widely available.
1.1 Interdisciplinary Context
Collaboration across several broad areas of research is critical, as different areas provide different information and context necessary to understanding the virus and disease.
This review provides a biological perspective on the virus Severe acute respiratory syndrome-related coronavirus 2 (SARS-CoV-2) and efforts to develop diagnostic, prophylactic, and therapeutic responses to COVID-19.
We provide only brief summaries of two other important perspectives on this pandemic: epidemiology and public health.
Research in these areas often seeks to anticipate, model, and prevent outbreaks of infectious disease or to understand and manage human behavior relevant to health and disease.
Their insights are critical to mounting a global response to the pandemic.
Epidemiological analyses have investigated patterns of transmission within and between communities, the symptoms associated with and the duration of infection and/or contagiousness, and how the virus propagates, among other characteristics [10].
Epidemiology also has a close relationship to public policy because it provides model-based insights into how preventative measures and public response can shift outcomes [11].
Public health addresses social and human factors influencing individuals’ exposure and susceptibility to pathogens, such as resource availability, inequality, human behavior, and access to accurate information.
Strategies from public health and epidemiology for managing the current epidemic have included the promotion of hand hygiene, social distancing, and personal protective equipment such as masks to mitigate spread, as well as containment approaches such as test, trace, and isolate, which depends on widespread testing, contact tracing, and quarantining.
An effective public health management strategy involves response coordination, disease surveillance, intervention monitoring, risk communication, and health education (including the containment of “infodemics” of false information) [12].
Epidemiology and public health intersect with the topics addressed in this manuscript because they both inform and benefit from relevant biotechnological developments.
For example, the development of accurate and fast diagnostic testing is relevant to test, trace, and isolate strategies for containment, and public education will be critical to deploying vaccines once they become available.
The present analysis focuses less on human and social factors and more on the basic biology of infection, diagnosis, and recovery, but these areas are inextricable in understanding and responding to the COVID-19 pandemic.
1.2 Initial Characterization of SARS-CoV-2
The first genome sequence of the virus was released on January 3, 2020 and revealed that the cluster of pneumonia cases seen in Wuhan were caused by a novel coronavirus [13].
Multiple research groups have drafted the genome sequence of SARS-CoV-2 based on sequences developed from clinical samples collected from the lower respiratory tract, namely bronchoalveolar lavage fluid (BALF), and the upper respiratory tract, in the form of throat swabs [14,15,16].
Analysis of the SARS-CoV-2 genome revealed significant sequence homology with two coronaviruses known to infect humans, with about 79% identity to SARS-CoV-1 and 50% to MERS-CoV [16].
However, the highest degree of similarity was observed between SARS-CoV-2 and bat-derived SARS-like coronaviruses (bat-SL-CoVZC45 and bat-SL-CoVZXC21) [15,16], with identity between SARS-CoV-2 and RATG13 as high as 96.2% [15,17].
This evidence therefore suggests the SARS-CoV-2 virus is the result of zoonotic transfer of a virus from bats to humans.
Nevertheless, some fragments between SARS-CoV-2 and RATG13 differ by up to 17%, suggesting a complex natural selection process during zoonotic transfer.
While the S region is highly similar to that of viruses found in pangolins [18], there is no consensus about the origin of S in SARS-CoV-2, as it could potentially be the result either of recombination or coevolution [17,19].
Though the intermediate host serving as the source for the zoonotic introduction of SARS-CoV-2 to human populations has not yet been identified, the SARS-CoV-2 virus has been placed within the coronavirus phylogeny through comparative genomic analyses.
Genomic analyses and comparisons to other known coronaviruses suggest that SARS-CoV-2 is unlikely to have originated in a laboratory – either purposely engineered and released, or escaped – and instead evolved naturally in an animal host [20].
While the position of the SARS-CoV-2 virus within the coronavirus phylogeny has been largely resolved, the functional consequences of molecular variation between this virus and other viruses, such as its bat and pangolin sister taxa or SARS-CoV-1, remain unknown [20].
Fortunately, the basic genome structure of coronaviruses is highly conserved, and insight into the mechanisms the virus uses to enter cells, replicate, and spread is likely to be available from prior research in coronaviruses.
1.3 Coronaviruses and Humans Hosts
Coronaviruses have long been known to infect animals and have been the subject of veterinary medical investigations and vaccine development efforts due to their effect on the health of companion and agricultural animals [21].
Most coronaviruses show little to no transmission in humans.
However, today it is thought that approximately one-third of common cold infections are caused by four human coronaviruses (HCoV): Human coronavirus 229E (HCoV-229E), Human coronavirus NL63 (HCoV-NL63), Human coronavirus OC43 (HCoV-OC43), and Human coronavirus HKU1 (HCoV-HKU1) [22,23].
The first HCoV were identified in the 1960s: HCoV-229E in 1965 [24] and HCoV-OC43 in 1967 [25].
Both of these viruses cause cold-like symptoms [26,27].
Two additional HCoV were subsequently identified [28,29].
In 2003, HCoV-NL63 [28] was first identified in a 7-month-old infant and then in clinical specimens collected from seven additional patients, five of whom were infants younger than 1 year old and the remainder of whom were adults.
CoV-HKU1 was identified in samples collected from a 71-year-old pneumonia patient in 2004 and then found in samples collected from a second adult patient [29].
These viruses are associated with respiratory diseases of varying severity, ranging from common cold to severe pneumonia, with severe symptoms mostly observed in immunocompromised individuals [30].
In addition to these relatively mild HCoV, however, highly pathogenic human coronaviruses have been identified, including Severe acute respiratory syndrome-related coronavirus (SARS-CoV or SARS-CoV-1) and Middle East respiratory syndrome-related coronavirus (MERS-CoV) [22,31,32].
At the time that SARS-CoV-1 emerged in the early 2000s, no HCoV had been identified in almost 40 years [31].
The first case of SARS was reported in November 2002 in the Guangdong Province of China, and over the following month, the disease spread more widely within China and then into several countries across multiple continents [31,33].
Unlike previously identified HCoV, SARS was much more severe, with an estimated death rate of 9.5% [33].
It was also highly contagious via droplet transmission, with a basic reproduction number (R0) of 4 (i.e., each person infected was estimated to infect four other people) [33].
However, the identity of the virus behind the infection remained unknown until April of 2003, when the SARS-CoV-1 virus was identified through a worldwide scientific effort spearheaded by the WHO [31].
SARS-CoV-1 belonged to a distinct lineage from the two other HCoV known at the time [33].
By July 2003, the SARS outbreak was officially determined to be under control, with the success credited to infection management practices [31].
A decade later, a second outbreak of severe respiratory illness associated with a coronavirus emerged, this time in the Arabian Peninsula.
This disease, known as Middle East respiratory syndrome (MERS), was linked to another novel coronavirus, MERS-CoV.
The fatality rate associated with MERS is much higher than that of SARS, at almost 35%, but the disease is much less easily transmitted, with an R0 of 1 [33].
Although MERS is still circulating, its low reproduction number has allowed for its spread to be contained [33].
The COVID-19 pandemic is thus associated with the seventh HCoV to be identified and the fifth since the turn of the millennium, though additional HCoVs may be in circulation but remain undetected.
SARS-CoV-1 and MERS-CoV were ultimately managed largely through infection management practices (e.g., mask wearing) and properties of the virus itself (i.e., low rate of transmission), respectively [31,33].
Vaccines were not used to control either virus, although vaccine development programs were established for SARS-CoV-1 [34].
In general, care for SARS and MERS patients focuses on supportive care and symptom management [33].
Clinical treatments for SARS and MERS developed during the outbreaks generally do not have strong evidence supporting their use.
Common treatments included Ribavirin, an antiviral, often in combination with corticosteroids or sometimes interferon (IFN) medications, which would both be expected to have immunomodulatory effects [31].
However, retrospective and in vitro analyses have reported inconclusive results of these treatments on SARS and the SARS-CoV-1 virus, respectively [31].
IFNs and Ribavirin have shown promise in in vitro analyses of MERS, but their clinical effectiveness remains unknown [31].
Therefore, only limited strategy for the pharmaceutical management of COVID-19 can be adopted from previous severe HCoV infections.
Research in response to prior outbreaks of HCoV-borne infections, such as SARS and MERS, have, however, provided a strong foundation for hypotheses about the pathogenesis of SARS-CoV-2 as well as potential diagnostic and therapeutic approaches.
1.3.1 Human Immune Response to Viral Threats
Understanding the fundamental organization of the human immune response to viral threats is critical to understanding the varied response to SARS-CoV-2.
The human immune system utilizes a variety of innate and adaptive responses to protect against the pathogens it encounters.
The innate immune system consists of barriers, such as the skin, mucous secretions, neutrophils, macrophages, and dendritic cells.
It also includes cell-surface receptors that can recognize the molecular patterns of pathogens.
The adaptive immune system utilizes antigen-specific receptors that are expressed on B and T lymphocytes.
These components of the immune system typically act together; the innate response acts first, and the adaptive response begins to act several days after initial infection following the clonal expansion of T and B cells [35].
After a virus enters into a host cell, its antigen is presented by major histocompatibility complex 1 (MHC 1) molecules and is then recognized by cytotoxic T lymphocytes.
In the case of COVID-19, there is also concern about the immune system becoming over-active.
One of the main immune responses contributing to the onset of acute respiratory distress syndrome (ARDS) in COVID-19 patients is cytokine storm syndrome (CSS), which causes an extreme inflammatory response due to a release of pro-inflammatory cytokines and chemokines by immune effector cells.
In addition to respiratory distress, this mechanism can lead to organ failure and death in severe COVID-19 cases [36].
Details of how the human body responds to SARS-CoV-2, both in healthy and pathological ways, and how these mechanisms can inform the identification of diagnostic, prophylactic, and therapeutic responses are explored in detail throughout this manuscript.
1.3.2 Clinical Presentation of COVID-19
A great diversity of symptom profiles has been observed for COVID-19, although a large study from Wuhan, China suggests fever and cough as the two most common symptoms on admission [37].
One early retrospective study in China described the clinical presentations of patients infected with SARS-CoV-2 as including lower respiratory tract infection with fever, dry cough, and dyspnea [38].
This study [38] noted that upper respiratory tract symptoms were less common, which suggests that the virus targets cells located in the lower respiratory tract.
However, data from the New York City region [39,40] showed variable rates of fever as a presenting symptom, suggesting that symptoms may not be consistent across samples.
These differences are present when comparing both between institutions in similar locations and between different regions experiencing COVID-19 outbreaks, leading to conflicting reports of the frequency of fever as a presenting symptom for patients upon hospital admission.
For example, even within New York City, one study [39] identified low oxygen saturation (<90% without the use of supplemental oxygen or ventilation support) in a significant percentage of patients upon presentation, while another study [40] reported cough, fever, and dyspnea as the most common presenting symptoms.
The variability of both which symptoms present and their severity makes it difficult for public health agencies to provide clear recommendations for citizens regarding what symptoms indicate SARS-CoV-2 infection and should prompt isolation.
1.4 Role of the COVID-19 Review
Several review articles on aspects of COVID-19 have already been published.
These have included reviews on the disease epidemiology [41], immunological response [42], diagnostics [43], and pharmacological treatments [42,44].
Others [45,46] provide narrative reviews of progress on some important ongoing COVID-19 research questions.
With the worldwide scientific community uniting during 2020 to investigate SARS-CoV-2 and COVID-19 from a wide range of perspectives, findings from many disciplines are relevant on a rapid timescale to a broad scientific audience.
Additionally, many findings are published as preprints, which are available prior to going through the peer review process.
As a result, centralizing, summarizing, and critiquing new literature broadly relevant to COVID-19 can help to expedite the interdisciplinary scientific process that is currently happening at an advanced pace.
We are particularly interested in providing background to the development of diagnostic, prophylactic, and therapeutic approaches to COVID-19.
Two major concerns within diagnosis include the detection of current infections in individuals with and without symptoms, and the detection of past exposure without an active infection.
In the latter category, identifying whether individuals can develop or have developed sustained immunity is also a major consideration.
The development of high-throughput, affordable methods for detecting active infections and sustained immunity will be critical to understanding and controlling the disease.
The identification of interventions that can mitigate the effect of the virus on exposed and infected individuals is a significant research priority.
Some possible approaches include the identification of existing pharmaceuticals that reduce the severity of infection, either by reducing the virus’ virulence (e.g., antivirals) or managing the most severe symptoms of infection.
Due to the long timeline for the development of novel pharmaceuticals, in most cases, research surrounding possible pharmaceutical interventions focuses on the identification and investigation of existing compounds whose mechanisms may be relevant to COVID-19.
Other foci of current research include the identification of antibodies produced by survivors of COVID-19 and the development of vaccines.
Understanding the mechanisms describing host-virus interactions between humans and SARS-CoV-2 is thus critical to identifying candidate therapeutics.
An overview of the topics covered is visualized in
Thus, in this review, we seek to consolidate information about efforts to develop strategies for diagnosis and therapeutics as new information is released by the scientific community.
We include information from both traditional peer-reviewed scientific literature and from preprints, which typically have not undergone peer review but have been critically evaluated by the scientists involved in this effort.
The goal of this manuscript is to present preliminary findings within the broader context of COVID-19 research and to identify the broad interpretations of new research, as well as limitations to interpretability.
2 Pathogenesis, Symptomatology, and Transmission of SARS-CoV-2 through analysis of Viral Genomics and Structure
2.1 Abstract
The novel coronavirus SARS-CoV-2, which emerged in late 2019, has since spread around the world infecting tens of millions of people with coronavirus disease 2019 (COVID-19).
While this viral species was unknown prior to January 2020, its similarity to other coronaviruses that infect humans has allowed for rapid insight into the mechanisms that it uses to infect human hosts, as well as the ways in which the human immune system can respond.
Here, we contextualize SARS-CoV-2 among other coronaviruses and identify what is known and what can be inferred about its behavior once inside a human host.
Because the genomic content of coronaviruses, which specifies the virus’s structure, is highly conserved, early genomic analysis provided a significant head start in predicting viral pathogenesis.
The pathogenesis of the virus offers insights into symptomatology, transmission, and individual susceptibility.
Additionally, prior research into interactions between the human immune system and coronaviruses has identified how these viruses can evade the immune system’s protective mechanisms.
We also explore systems-level research into the regulatory and proteomic effects of SARS-CoV-2 infection and the immune response.
Understanding the structure and behavior of the virus serves to contextualize the many facets of the COVID-19 pandemic and can influence efforts to control the virus and treat the disease.
2.2 Importance
COVID-19 involves a number of organ systems and can present with a wide range of symptoms.
Understanding how the virus infects epithelial cells, however, serves to contextualize how these systems connect.
Similarly, while the modes of viral transmission have been under debate throughout much of 2020 and the beginning of 2021, the available research suggests that these patterns are very similar to those seen in the closely related viruses SARS-CoV-1 and possibly MERS-CoV.
Exploring the structure, phylogeny, and pathogenesis of the virus therefore helps to guide interpretation of the broader impacts of the virus on the human body and on human populations.
For this reason, an in-depth exploration of viral mechanisms is critical to a robust understanding of the COVID-19 pandemic.
2.3 Introduction
The current coronavirus disease 2019 (COVID-19) pandemic, caused by the Severe acute respiratory syndrome-related coronavirus 2 (SARS-CoV-2) virus, represents an acute global health crisis.
Symptoms of the disease can range from mild to severe or fatal [47] and can affect a variety of organs and systems [48].
Outcomes of infection can include acute respiratory distress (ARDS) and acute lung injury, as well as damage to other organ systems [48,49].
Understanding the progression of the disease, including these diverse symptoms, depends on understanding how the virus interacts with the host.
Additionally, the fundamental biology of the virus can provide insights into how it is transmitted among people, which can, in turn, inform efforts to control its spread.
As a result, a thorough understanding of the pathogenesis of SARS-CoV-2 is a critical foundation on which to build an understanding of COVID-19 and the pandemic as a whole.
The rapid identification and release of the genomic sequence of the virus in January 2020 [13] provided early insight into the virus in a comparative genomic context.
The viral genomic sequence clusters with known coronaviruses (order Nidovirales, family Coronaviridae, subfamily Orthocoronavirinae).
Phylogenetic analysis of the coronaviruses reveals four major subclades, each corresponding to a genus: the alpha, beta, gamma, and delta coronaviruses.
Among them, alpha- and betacoronaviruses infect mammalian species, gammacoronaviruses infect avian species, and deltacoronaviruses infect both mammalian and avian species [50].
The novel virus now known as SARS-CoV-2 was identified as a betacoronavirus belonging to the B lineage based on phylogenetic analysis of a polymerase chain reaction (PCR) amplicon fragment from five patients along with the full genomic sequence [51].
This lineage also includes the Severe acute respiratory syndrome-related coronavirus (SARS-CoV-1) that caused the 2002-2003 outbreak of Severe Acute Respiratory Syndrome (SARS) in humans [51].
Because viral structure and mechanisms of pathogenicity are highly conserved within the order, this phylogenetic analysis provided a basis for forming hypotheses about how the virus interacts with hosts, including which tissues, organs, and systems would be most susceptible to SARS-CoV-2 infection.
Coronaviruses that infect humans (HCoV) are not common, but prior research into other HCoV such as SARS-CoV-1 and Middle East respiratory syndrome-related coronavirus (MERS-CoV), as well as other viruses infecting humans such as a variety of influenza species, established a strong foundation that accelerated the pace of SARS-CoV-2 research.
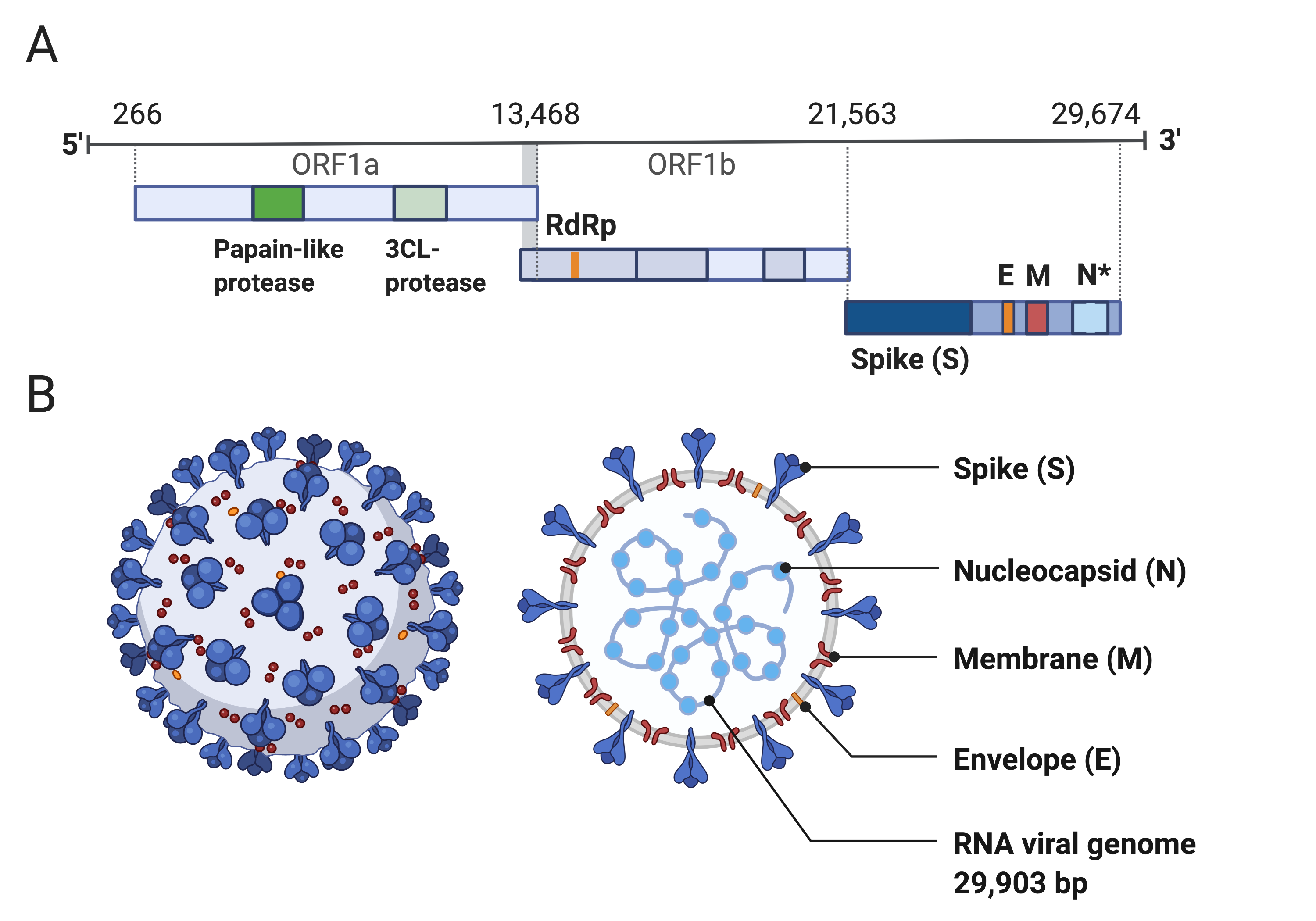
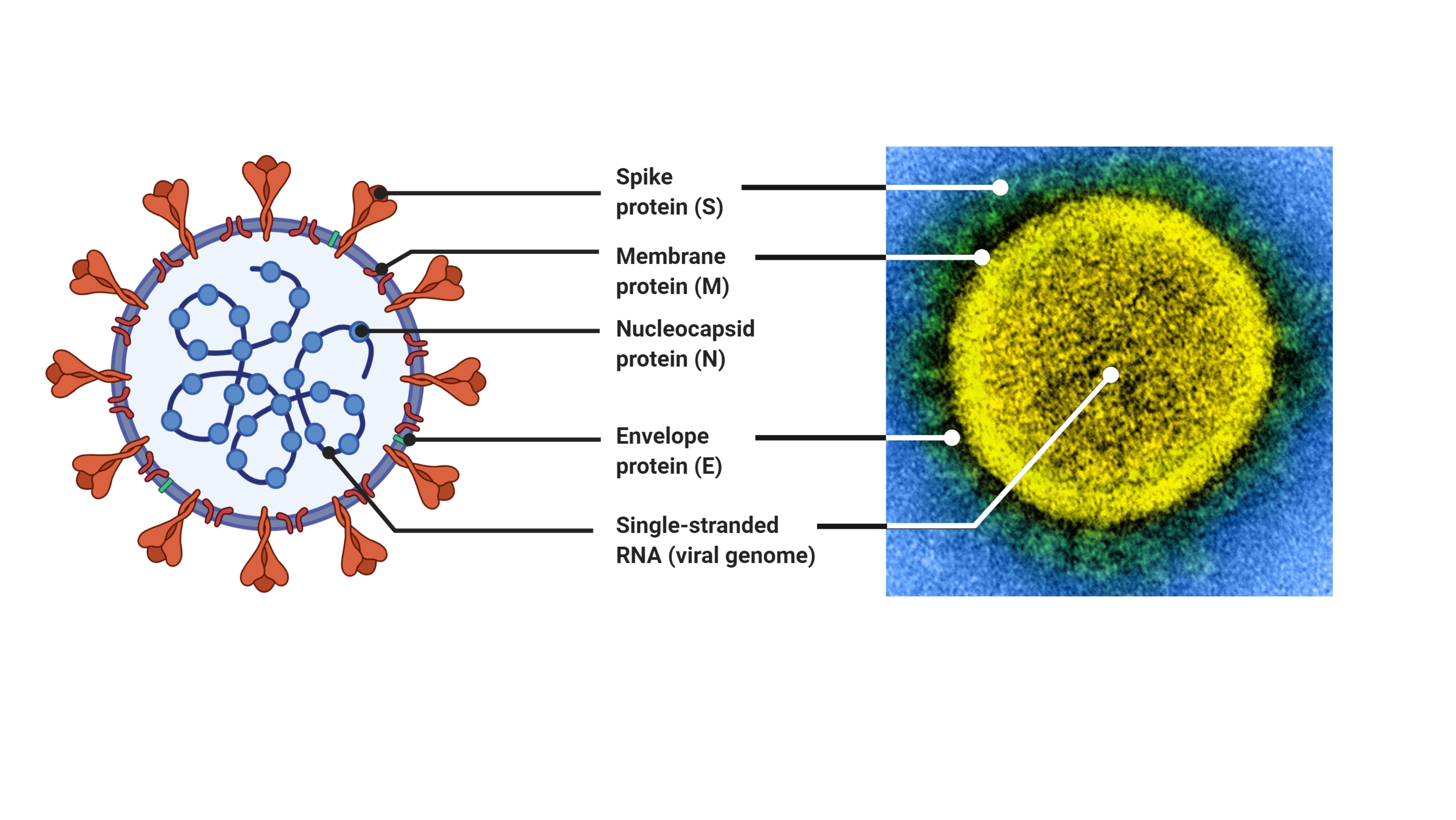
Coronaviruses are large viruses that can be identified by their distinctive “crown-like” shape (Figure 1).
Their spherical virions are made from lipid envelopes ranging from 100 to 160 nanometers in which peplomers (protruding structures) of two to three spike (S) glycoproteins are anchored, creating the crown [52,53].
These spikes, which are critical to both viral pathogenesis and to the response by the host immune response, have been visualized using cryo-electron microscopy [54].
Because they induce the human immune response, they are also the target of many proposed therapeutic agents.
Viral pathogenesis is typically broken down into three major components: entry, replication, and spread [55].
However, in order to draw a more complete picture of pathogenesis, it is also necessary to examine how infection manifests clinically, identify systems-level interactions between the virus and the human body, and consider the possible effects of variation or evolutionary change on pathogenesis and virulence.
Thus, clinical medicine and traditional biology are both important pieces of the puzzle of SARS-CoV-2 presentation and pathogenesis.
2.4 Coronavirus Structure and Pathogenesis
2.4.1 Structure of Coronaviruses
Genome structure is highly conserved among coronaviruses, meaning that the relationship between the SARS-CoV-2 genome and its pathogenesis can be inferred from prior research in related viral species.
The genomes of viruses in the Nidovirales order share several fundamental characteristics.
They are non-segmented, which means the viral genome is contained in a single capsid, and are enveloped, which means that the genome and capsid are encased by a lipid bilayer.
Coronaviruses have large positive-sense RNA (ssRNA+) genomes ranging from 27 to 32 kilobases in length [16,56].
The SARS-CoV-2 genome lies in the middle of this range at 29,903 bp [16].
Genome organization is highly conserved within the order [56].
There are three major genomic regions: one containing the replicase gene, one containing the genes encoding structural proteins, and interspersed accessory genes [56] (Figure 1).
The replicase gene comprises about two-thirds of the genome and consists of two open reading frames that are translated with ribosomal frameshifting [56].
This polypeptide is then translated into 16 non-structural proteins (nsp), except in gammacoronaviruses where nsp1 is absent, that form the replication machinery used to synthesize viral RNA [57].
The remaining third of the genome encodes structural proteins, including the spike, membrane, envelope, and nucleocapsid proteins.
Additional accessory genes are sometimes present between these two regions, depending on the species or strain.
Much attention has been focused on the S protein, which is a critical structure involved in cell entry.
Figure 1:Structure of SARS-CoV-2 capsid and genome.
A) The genomic structure of coronaviruses is highly conserved and includes three main regions.
Open reading frames (ORF) 1a and 1b contain two polyproteins that encode the non-structural proteins (nsp).
The nsp include proteases such as RNA-dependent RNA Polymerase (RdRp).
The last third of the genome encodes structural proteins, including the spike (S), envelope (E), membrane (M) and nucleocapsid (N) proteins.
Accessory genes can also be interspersed throughout the genome [56].
B) The physical structure of the coronavirus virion, including the components determined by the conserved structural proteins S, E, M and N.
2.4.2 Pathogenic Mechanisms of Coronaviruses
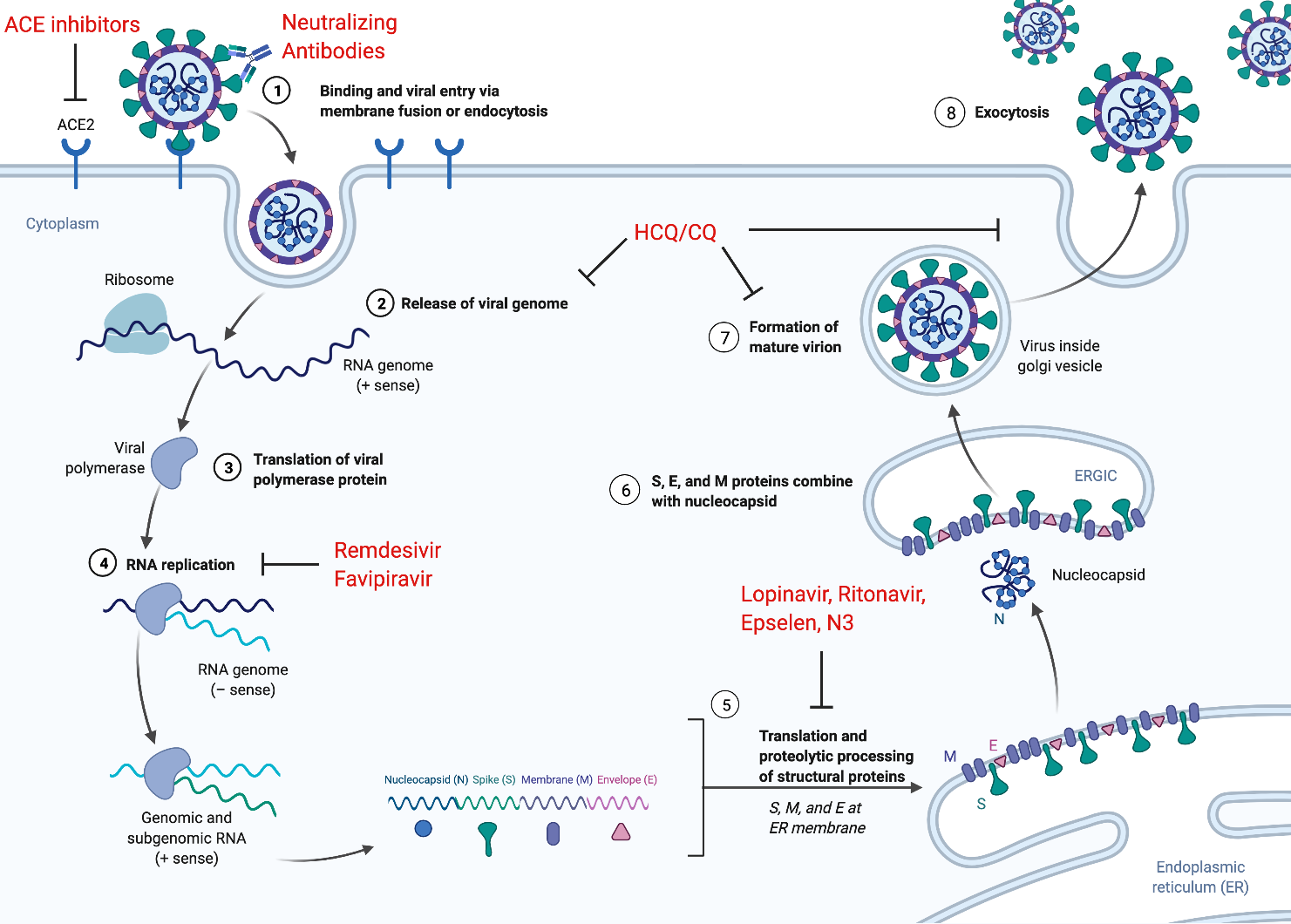
While, like most viruses, it is possible that SARS-CoV-1 and SARS-CoV-2 can enter cells through endocytosis, a process conserved among coronaviruses enables them to target cells for entry through fusion with the plasma membrane [58,59].
Cell entry proceeds in three steps: binding, cleavage, and fusion.
First, the viral spike protein binds to a host cell via a recognized receptor or entry point.
Coronaviruses can bind to a range of host receptors [60,61], with binding conserved only at the genus level [50].
Viruses in the betacoronavirus genus, to which SARS-CoV-2 belongs, are known to bind to the CEACAM1 protein, 5-N-acetyl-9-O-acetyl neuraminic acid, and to the angiotensin-converting enzyme 2 (ACE2) [60].
SARS-CoV-2 has a high affinity for human ACE2, which is expressed in the vascular epithelium, other epithelial cells, and cardiovascular and renal tissues [62,63], as well as many others [64].
The binding process is guided by the molecular structure of the spike protein, which is structured in three segments: an ectodomain, a transmembrane anchor, and an intracellular tail [65].
The ectodomain forms the crown-like structures on the viral membrane and contains two subdomains known as the S1 and S2 subunits [66].
The S1 (N-terminal) domain forms the head of the crown and contains the receptor binding motif, and the S2 (C-terminal) domain forms the stalk that supports the head [66].
The S1 subunit guides the binding of the virus to the host cell, and the S2 subunit guides the fusion process [65].
After the binding of the S1 subunit to an entry point, the spike protein is often cleaved at the S1-S2 boundary by a host protease [67,68,69].
Similar to SARS-CoV-1, SARS-CoV-2 exhibits redundancy in which host proteases can cleave the S protein [70].
Specifically, both transmembrane protease serine protease-2 (TMPRSS2) and cathepsins B/L have been shown to mediate SARS-CoV-2 S protein proteolytic priming, and small molecule inhibition of these enzymes fully inhibited viral entry in vitro[70,71].
Proteolytic priming prepares the S protein for fusion [68,69].
The two subunits remain bound by van der Waals forces, with the S1 subunit stabilizing the S2 subunit during the membrane fusion process [67].
Electron microscopy suggests that in some coronaviruses, including SARS-CoV-1 and MERS-CoV, a six-helix bundle separates the two subunits in the postfusion conformation, and the unusual length of this bundle facilitates membrane fusion through the release of additional energy [50].
Cleavage at a second site within S2 by these same proteases activates S for fusion by inducing conformational changes [67].
The viral membrane can then fuse with the endosomal membrane to release the viral genome into the host cytoplasm.
Once the virus enters a host cell, the replicase gene is translated and assembled into the viral replicase complex.
This complex then synthesizes the double-stranded RNA (dsRNA) genome from the genomic ssRNA(+).
The dsRNA genome is transcribed and replicated to create viral mRNAs and new ssRNA(+) genomes [56,72].
From there, the virus can spread into other cells.
In this way, the genome of SARS-CoV-2 provides insight into the pathogenic behavior of the virus.
Evidence also suggests that SARS-CoV-2 may take advantage of the specific structure of endothelial cells to enter the circulatory system.
Endothelial cells are specialized epithelial cells [73] that form a barrier between the bloodstream and surrounding tissues.
The endothelium facilitates nutrient, oxygen, and cellular exchange between the blood and vascularized tissues [74].
The luminal (interior) surface of the endothelium is lined with glycocalyx, a network of both membrane-bound and soluble proteins and carbohydrates, primarily proteoglycans and glycoproteins [75,76].
The glycocalyx varies in thickness from 0.5 microns in the capillaries to 4.5 microns in the carotid arteries and forms a meshwork that localizes both endothelial- and plasma-derived signals to the inner vessel wall [75].
Heparan sulfate is the dominant proteoglycan in the glycocalyx, representing 50-90% of glycocalyx proteoglycan content [77].
The SARS-CoV-2 spike protein can bind directly to heparan sulfate, which serves in part as a scaffolding molecule to facilitate ACE2 binding and entry into endothelial cells [76].
A heparan sulfate binding site has also been identified near the ACE2 binding site on the viral RBD, and modeling has suggested that heparan sulfate binding yields an open conformation that facilitates binding to ACE2 on the cell surface [76].
Degrading or removing heparan sulfate was associated with decreased binding [76].
Heparan sulfate may also interact with the S1/S2 proteolytic cleavage site and other binding sites to promote binding affinity [78].
Notably, treatment with soluble heparan sulfate or even heparin (a commonly used anti-coagulant and vasodilator that is similar in structure to heparan sulfate [79]) potently blocked spike protein binding and viral infection [76].
This finding is particularly interesting because degradation of heparan sulfate in the glycocalyx has previously been identified as an important contributor to ARDS and sepsis [80], two common and severe outcomes of COVID-19, and suggests that heparan sulfate could be a target for pharmaceutical inhibition of cell entry by SARS-CoV-2 [81,82,83,84,85].
Together, this evidence suggests that heparan sulfate can serve as an important adhesion molecule for SARS-CoV-2 cell entry and may represent a therapeutic target.
2.4.3 Immune Evasion Strategies
Research in other HCoV provides some indication of how SARS-CoV-2 infection proceeds in spite of the human immune response.
By infecting the epithelium, viruses such as SARS-CoV-1 are known to bypass the physical barriers, such as skin and mucus, that comprise the immune system’s first line of defense [86].
Once the virus infiltrates host cells, it is adept at evading detection.
CD163+ and CD68+ macrophage cells are especially crucial for the establishment of SARS-CoV-1 in the body [86].
These cells most likely serve as viral reservoirs that help shield SARS-CoV-1 from the innate immune response.
According to a study on the viral dissemination of SARS-CoV-1 in Chinese macaques, viral RNA could be detected in some monocytes throughout the process of differentiation into dendritic cells [86].
This lack of active viral replication allows SARS-CoV-1 to escape the innate immune response because reduced levels of detectable viral RNA allow the virus to avoid both natural killer cells and Toll-like receptors [86].
Even during replication, SARS-CoV-1 is able to mask its dsRNA genome from detection by the immune system.
Although dsRNA is a pathogen-associated molecular pattern that would typically initiate a response from the innate immune system [87], in vitro analysis of nidoviruses including SARS-CoV-1 suggests that these viruses can induce the development of double-membrane vesicles that protect the dsRNA signature from being detected by the host immune system [88].
This protective envelope can therefore insulate these coronaviruses from the innate immune system’s detection mechanism [36].
HCoVs are also known to interfere with the host immune response, rather than just evade it.
For example, the virulence of SARS-CoV-2 is increased by nsp1, which can suppress host gene expression by stalling mRNA translation and inducing endonucleolytic cleavage and mRNA degradation [89].
SARS-CoV-1 also evades the immune response by interfering with type I IFN induction signaling, which is a mechanism that leads to cellular resistance to viral infections.
SARS-CoV-1 employs methods such as ubiquitination and degradation of RNA sensor adaptor molecules MAVS and TRAF3/6 [90].
Also, MERS-CoV downregulates antigen presentation via MHC class I and MHC class II, which leads to a reduction in T cell activation [90].
These evasion mechanisms, in turn, may facilitate systemic infection.
Coronaviruses such as SARS-CoV-1 are also able to evade the humoral immune response through other mechanisms, such as inhibiting certain cytokine pathways or down-regulating antigen presentation by the cells [88].
2.4.4 Host Cell Susceptibility
ACE2 and TMPRSS2 have been identified as the primary entry portal and as a critical protease, respectively, in facilitating the entry of SARS-CoV-1 and SARS-CoV-2 into a target cell [54,70,91,92,93].
This finding has led to a hypothesized role for ACE2 and TMPRSS2 expression in determining which cells, tissues, and organs are most likely to be infected by SARS-CoV-2.
ACE2 is expressed in numerous organs, such as the heart, kidney, and intestine, but it is most prominently expressed in alveolar epithelial cells; this pattern of expression is expected to contribute to the virus’ association with lung pathology [62,94,95] as well as that of SARS [96].
Clinical investigations of COVID-19 patients have detected SARS-CoV-2 transcripts in bronchoalveolar lavage fluid (BALF) (93% of specimens), sputum (72%), nasal swabs (63%), fibrobronchoscopy brush biopsies (46%), pharyngeal swabs (32%), feces (29%), and blood (1%) [97].
Two studies reported that SARS-CoV-2 could not be detected in urine specimens [97,98]; however, a third study identified four urine samples (out of 58) that were positive for SARS-CoV-2 nucleic acids [99].
Although respiratory failure remains the leading cause of death for COVID-19 patients [100], SARS-CoV-2 infection can damage many other organ systems including the heart [101], kidneys [102,103], liver [104], and gastrointestinal tract [105,106].
As it becomes clear that SARS-CoV-2 infection can damage multiple organs, the scientific community is pursuing multiple avenues of investigation in order to build a consensus about how the virus affects the human body.
2.5 Clinical Presentation of COVID-19
SARS-CoV-2 pathogenesis is closely linked with the clinical presentation of the COVID-19 disease.
Reports have described diverse symptom profiles associated with COVID-19, with a great deal of variability both within and between institutions and regions.
A large study from Wuhan, China conducted early in the pandemic identified fever and cough as the two most common symptoms that patients reported at hospital admission [37], while a retrospective study in China described the clinical presentations of patients infected with SARS-CoV-2 as including lower respiratory tract infection with fever, dry cough, and dyspnea (shortness of breath) [38].
This study [38] noted that upper respiratory tract symptoms were less common, suggesting that the virus preferentially targets cells located in the lower respiratory tract.
However, data from the New York City region [39,40] showed variable rates of fever as a presenting symptom, suggesting that symptoms may not be consistent across individuals.
For example, even within New York City, one study [39] identified low oxygen saturation (<90% without the use of supplemental oxygen or ventilation support) in 20.4% of patients upon presentation, with fever being present in 30.7%, while another study [51] reported cough (79.4%), fever (77.1%), and dyspnea (56.5%) as the as the most common presenting symptoms; both of these studies considered only hospitalized patients.
A later study reported radiographic findings such as ground-glass opacity and bilateral patchy shadowing in the lungs of many hospitalized patients, with most COVID-19 patients having lymphocytopenia, or low levels of lymphocytes (a type of white blood cell) [37].
Patients may also experience loss of smell, myalgias (muscle aches), fatigue, or headache.
Gastrointestinal symptoms can also present [107], and the CDC includes nausea and vomiting, as well congestion and runny nose, on its list of symptoms consistent with COVID-19 [47].
A preprint using data from an app-based survey of 500,000 individuals in the US found that among those tested for SARS-CoV-2, a loss of taste or smell, fever, and a cough were significant predictors of a positive test result [108].
It is important to note that in this study, the predictive value of symptoms may be underestimated if they are not specific to COVID-19.
This underestimation could occur because the outcome measured was a positive, as opposed to a negative, COVID-19 test result, meaning an association would be more easily identified for symptoms that were primarily or exclusively found with COVID-19.
At the time the surveys were conducted, due to limits in US testing infrastructure, respondents typically needed to have some symptoms known to be specific to COVID-19 in order to qualify for testing.
Widespread testing of asymptomatic individuals may therefore provide additional insight into the range of symptoms associated with COVID-19.
Consistent with the wide range of symptoms observed and the pathogenic mechanisms described above, COVID-19 can affect diverse body systems in addition to causing respiratory problems [109].
For example, COVID-19 can lead to acute kidney injury, especially in patients with severe respiratory symptoms or certain preexisting conditions [110].
It can also cause neurological complications [111,112,113], potentially including stroke, seizures or meningitis [114,115].
In fact, autopsy samples suggest that SARS-CoV-2 may be able to enter the central nervous system via the neural–mucosal interface [116].
COVID-19 has also been associated with an increased incidence of large vessel stroke, particularly in patients under the age of 40 [117], and other thrombotic events including pulmonary embolism and deep vein thrombosis [118].
The mechanism behind these complications has been suggested to be related to coagulopathy, with reports indicating the presence of antiphospholipid antibodies [119] and elevated levels of d-dimer and fibrinogen degradation products in deceased patients [120].
Other viral infections have been associated with coagulation defects and changes to the coagulation cascade; notably, SARS was also found to lead to disseminated intravascular coagulation and was associated with both pulmonary embolism and deep vein thrombosis [121].
The mechanism behind these insults has been suggested to be related to inflammation-induced increases in the von Willebrand factor clotting protein, leading to a pro-coagulative state [121].
Abnormal clotting (thromboinflammation or coagulopathy) has been increasingly discussed recently as a possible key mechanism in many cases of severe COVID-19, and may be associated with the high d-dimer levels often observed in severe cases [122,123,124].
This excessive clotting in lung capillaries has been suggested to be related to a dysregulated activation of the complement system, part of the innate immune system [125,126].
2.5.1 Pediatric Presentation
The presentation of COVID-19 infection can vary greatly among pediatric patients and, in some cases, manifests in distinct ways from COVID-19 in adults.
Evidence suggests that while children and adolescents tend to have mostly asymptomatic infections, those that are symptomatic typically exhibit a mild illness [127,128,129,130].
One review examined symptoms reported in 17 studies of children infected with COVID-19 during the early months of the COVID-19 epidemic in China and one study from Singapore [131].
In the more than a thousand cases described, the most common reports were for mild symptoms such as fever, dry cough, fatigue, nasal congestion and/or runny nose, while three children were reported to be asymptomatic.
Severe lower respiratory infection was described in only one of the pediatric cases reviewed.
Gastrointestinal symptoms such as vomiting or diarrhea were occasionally reported.
Radiologic findings were not always reported in the case studies reviewed, but when they were mentioned they included bronchial thickening, ground-glass opacities, and/or inflammatory lesions [131].
Neurological symptoms have also been reported [132].
These analyses indicate that most pediatric cases of COVID-19 are not severe.
Indeed, it is estimated that less than 1% of pediatric cases result in critical illness [129,133].
However, serious complications and, in rare cases, deaths have occurred [134].
Of particular interest, children have occasionally experienced a serious inflammatory syndrome, multisystem inflammatory syndrome in children (MIS-C), following COVID-19 infection.
This syndrome is similar in some respects to Kawasaki disease, including Kawasaki disease shock syndrome [135,136,137] and is thought to be a distinct clinical manifestation of SARS-CoV-2 due to its distinct cytokine profile and the presence of burr cells in peripheral blood smears [138,139].
MIS-C has been associated with heart failure in some cases [140].
One case study [141] described an adult who appeared to show symptoms similar to MIS-C after exposure to COVID-19, but cautioned against broad conclusions; a second possible adult case has also been reported [142].
The presentation of SARS-CoV-2 infection is therefore likely to be largely distinct between adult and pediatric populations.
However, not all cases of severe COVID-19 in children are characterizable as MIS-C.
A recent study [143] described demographic and clinical variables associated with MIS-C in comparison with non-MIS-C severe acute COVID-19 in young people in the United States.
2.5.2 Cytokine Release Syndrome
Symptoms of a disease can be caused by a pathogen, but they can also be caused by the immune system’s reaction to the pathogen.
A dysregulated immune response can cause significant damage to the host [144,145,146].
The inflammatory response has received particular attention for its role in both a healthy response to infection and a pathogenic one.
Inflammation is one of the most visible components of the immune response, as it is responsible for the hallmarks of injury, such as pain, heat, and swelling [147].
In response to injury or to signaling by pattern recognition receptors indicating the detection of a molecular pattern associated with a pathogen or foreign body, the immune system stimulates leukocytes that travel to the site of the threat, where they then produce cytokines [147].
Cytokines are a diverse group of small proteins that play an important role in intercellular signaling [148].
Cytokines can be both pro- and anti-inflammatory, which means they can either stimulate or inhibit the production of additional cytokines [148,149].
Some notable pro-inflammatory cytokines include the interleukins IL-1β and IL-6 and tumor necrosis factor α (TNF-α) [149].
Anti-inflammatory cytokines play an immunoregulatory role complementary to the cascading effect of pro-inflammatory cytokines [148,149].
A number of interleukins and interferons play anti-inflammatory roles, and receptors or receptor antagonists for inflammatory cytokines are also important for regulating inflammation [149].
IL-10 is an anti-inflammatory cytokine of particular note because it regulates the expression of TNF-α, IL-1, and IL-6 [149].
When the pro- and anti-inflammatory responses are both commensurate with the threat posed, the immune system drives a shift back to homeostasis [150].
However, when the responses are disproportionate, the cytokine response can become dysregulated.
Too low of an inflammatory response will not eliminate the immune threat [150].
In contrast, if the response is dysregulated towards excessive pro-inflammatory cytokine activity, inflammation can cascade [151] and cause cell damage, among other problems [147].
Elevated levels of inflammation over the long-term are associated with many chronic health conditions, including type 2 diabetes, dementia and Alzheimer’s, and arthritis [152].
On a shorter timescale, dysregulated systemic inflammation can cause sepsis, which can lead to multi-organ failure and death [148,153].
Cytokines have been investigated for their role in the immune response to lung infections long before the COVID-19 pandemic.
Dysregulation of the inflammatory response, including elevated levels of pro-inflammatory cytokines, is found in patients with ARDS, which is a severe condition that can arise from pneumonia, SARS, and COVID-19 [151].
One study of patients with and at risk for ARDS, specifically those who were intubated for medical ventilation, found that shortly after the onset of ARDS, anti-inflammatory cytokine concentration in BALF increased relative to the concentration of pro-inflammatory cytokines [154].
The results suggest that an increase in pro-inflammatory cytokines such as IL-6 may signal the onset of ARDS, but recovery depends on an increased anti-inflammatory response [154].
However, patients with severe ARDS were excluded from this study.
Acute phase response to an infection can also cause damage to the capillary endothelium, allowing leaks that disrupt the balance between pro-inflammatory cytokines and their regulators [154].
Hyperactivity of the pro-inflammatory response due to lung infection is commonly associated with acute lung injury and more rarely with the more severe manifestation, ARDS [148].
The heightened inflammatory response in the lungs can also serve as a source for systemic inflammation, or sepsis, and potentially multi-organ failure [148].
The shift from local to systemic inflammation is a phenomenon often referred to broadly as a cytokine storm [148] or, more precisely, as cytokine release syndrome [155].
Sepsis is a known possible complication of pneumonia, and in an analysis of over 1,400 US pneumonia patients, IL-6, tumor necrosis factor (TNF), and IL-10 were found to be elevated at intake in patients who developed severe sepsis and/or ultimately deceased [156].
However, unlike the study analyzing pro- and anti-inflammatory cytokines in ARDS patients [154], this study reported that unbalanced pro-/anti-inflammatory cytokine profiles were rare.
This discrepancy could be related to the fact that the sepsis study measured only three cytokines.
Regardless of variation in the anti-inflammatory response, prior work has therefore made it clear that pulmonary infection and injury are associated with systemic inflammation and with sepsis.
While IL-6 is a biomarker sometimes used to assess cytokine storm activity in sepsis [148], the relationship between cytokine profiles and the risks associated with sepsis may be more complex.
In fact, although IL-6 has traditionally been considered pro-inflammatory, its pleiotropic effects via both classical and trans-signaling allow it to play an integral role in both the inflammatory and anti-inflammatory responses [157], leading it to be associated with both healthy and pathological responses to viral threat [158].
The inflammatory response was identified early on as a potential driver of COVID-19 outcomes due to existing research in SARS and emerging research in COVID-19.
In addition to the known role of cytokines in ARDS and lung infection more broadly, immunohistological analysis at autopsy of patients deceased from SARS revealed that ACE2-expressing cells that were infected by SARS-CoV-1 showed elevated expression of IL-6, IL-1β, and TNF-α [159].
Similarly, the introduction of the S protein from SARS-CoV-1 to mouse macrophages was found to increase production of IL-6 and TNF-α [160].
For SARS-CoV-2 infection leading to COVID-19, early reports described a cytokine storm syndrome-like response in patients with particularly severe infections [94,161,162].
Among patients hospitalized with COVID-19 in Wuhan, China, 112 out of 191 (59%) developed sepsis, including all 54 of the non-survivors [38].
However, the argument has been made that while the cytokine levels observed in COVID-19 patients fall outside of the normal range, they are not as high as typically found in patients with ARDS [163].
Regardless, inflammation has received significant interest both in regards to the pathology of COVID-19 as well as potential avenues for treatment, as the relationship between the cytokine storm and the pathophysiology of COVID-19 has led to the suggestion that a number of immunomodulatory pharmaceutical interventions could hold therapeutic value for the treatment of COVID-19 [164].
2.6 Systems-Level Effects
Systems biology provides a cross-disciplinary analytical paradigm through which the host response to an infection can be analyzed.
This field integrates the “omics” fields (genomics, transcriptomics, proteomics, metabolomics, etc.) using bioinformatics and other computational approaches.
Over the last decade, systems biology approaches have been used widely to study the pathogenesis of diverse types of life-threatening acute and chronic infectious diseases [165].
Omics-based studies have also provided meaningful information regarding host immune responses and surrogate protein markers in several viral, bacterial and protozoan infections [166].
Though the complex pathogenesis and clinical manifestations of SARS-CoV-2 infection are not yet fully understood, omics technologies offer the opportunity for discovery-driven analysis of biological changes associated with SARS-CoV-2 infection.
For example, previous studies suggest that infection by coronaviruses, such as SARS-CoV-1 and MERS-CoV, as well as other viruses, is associated with the upregulation of ACE2.
In several preliminary assays and an analysis of microarray data, ACE2 expression was reported to be significantly upregulated following infection of human embryonic kidney cells and human airway epithelial cells [94].
This study also reported that direct stimulation with inflammatory cytokines such as type I interferons (e.g., IFNβ) resulted in the upregulation of ACE2 in human bronchial epithelial cells, with treated groups showing four-fold higher ACE2 expression than control groups at 18 hours post-treatment [94].
While it is still unclear whether SARS-CoV-2 facilitates the positive regulation of its own transmission between host cells, the host immune response itself likely plays a key role in mediating infection-associated pathologies.
For this reason, the application of omics technologies to the process of characterizing the host response is expected to provide novel insights into how hosts respond to SARS-CoV-2 infection and how these changes might influence COVID-19 outcomes.
2.6.1 Transcriptomics
In addition to the study described above [94], two other studies have profiled expression following SARS-CoV-2 infection using human cell lines.
The first study [167] compared transcriptional responses to SARS-CoV-2 and to other respiratory viruses, including MERS-CoV, SARS-CoV, Human parainfluenza virus 3, Respiratory syncytial virus, and Influenza A virus.
The responses of three human cell lines were analyzed: A549 (adenocarcinomic human alveolar basal epithelial cells), Calu-3 (human airway epithelial cells derived from human bronchial submucosal glands), and MRC-5 (human fetal lung fibroblast cells).
As the viral entry portal ACE2 has low expression in A549 cells, these cells were supplemented with adenovirus-based vectors expressing either mCherry (a fluorescent protein used as a control) or ACE2 (A549-ACE2).
The authors also measured host transcriptional responses to SARS-CoV-2 in primary normal human bronchial epithelial cells (HBEC or NHBE cells), nasal washes from an animal model (ferret), and lung samples from two COVID-19 patients.
The transcriptional response differed between the COVID-19 infected cells and the cells infected by other viruses, with changes in differential expression specific to each infection type.
In the hosts where SARS-CoV-2 was able to replicate efficiently, differential expression analysis revealed that the transcriptional response was significantly different from the response to all of the other viruses tested.
A unique pro-inflammatory cytokine signature associated with SARS-CoV-2 was present in cells exposed to both high and low doses of the virus, with the cytokines IL-6 and IL1RA uniquely elevated in response to SARS-CoV-2 relative to other viruses.
However, the A549-ACE2 cells showed significant IFN-I or IFN-III expression when exposed to high, but not low, doses of SARS-CoV-2.
This finding suggests that IFN induction is dependent on the extent of exposure.
Similarly, in cells from the NHBE line, ferrets, and COVID-19 patients, chemokine signaling was significantly enriched, but there was no significant induction of IFN-I or IFN-III.
Together, these results suggest that SARS-CoV-2 induces a limited antiviral state with low IFN-I or IFN-III expression and a moderate IFN-stimulated gene response, in contrast to other viruses.
Other respiratory viruses have been found to encode antagonists to the IFN response.
The analysis of SARS-CoV-2 suggested that this transcriptional state was specific to cells expressing ACE2, as it was not observed in cells lacking expression of this protein except with ACE2 supplementation and at very high (10-fold increase) level of SARS-CoV-2 exposure.
This hypothesis was further supported by a recent study [168] that showed that the SARS-CoV-2 ORF3b gene suppresses IFNB1 promoter activity (IFN-I induction) more efficiently than the SARS-CoV-1 ORF3b gene.
Taken together, these findings suggest that a unique cytokine profile is associated with the response to the SARS-CoV-2 virus, and that this response differs depending on the magnitude of exposure.
Another study [169] analyzed dynamic transcriptional responses to SARS-CoV-2 and SARS-CoV-1.
They characterized the response of three human cell lines, H1299 (human non-small cell lung carcinoma cell line), Calu-3, and Caco-2 (human epithelial colorectal adenocarcinoma cell line), at 4 to 36 hours post infection.
Using poly(A) bulk RNA-seq, the authors found negligible susceptibility of H1299 cells (< 0.08 viral read percentage of total reads) compared to Caco-2 and Calu-3 cells (>10% of viral reads).
This finding suggests that the risk of infection varies among cell types, and that cell type could influence which hosts are more or less susceptible.
Based on visual inspection of microscopy images alongside transcriptional profiling, the authors also showed distinct responses among the host cell lines evaluated.
In contrast to Caco-2, Calu-3 cells infected with SARS-CoV-2 showed signs of impaired growth and cell death at 24 hours post infection, as well as moderate IFN induction with a strong up-regulation of IFN-stimulated genes.
Interestingly, the results were similar to those reported in Calu-3 cells exposed to much higher levels of SARS-CoV-2 [167], as described above.
This finding suggests that IFN induction in Calu-3 cells is not dependent on the level of exposure, in contrast to A549-ACE2 cells.
The discrepancy could be explained by the observations that Calu-3 cells are highly susceptible to SARS-CoV-2 and show rapid viral replication [71], whereas A549 cells are incompatible with SARS-CoV-2 infection [170].
This discrepancy raises the concern that in vitro models may vary in their similarity to the human response, underscoring the importance of follow-up studies in additional models.
2.6.2 Proteomics
One early proteomics study investigated changes associated with in vitro SARS-CoV-2 infection using Caco-2 cells [171].
This study reported that SARS-CoV-2 induced alterations in multiple vital physiological pathways, including translation, splicing, carbon metabolism and nucleic acid metabolism in the host cells.
Another area of interest is whether SARS-CoV-2 is likely to induce similar changes to other HCoV.
For example, because of the high level of sequence homology between SARS-CoV-2 and SARS-CoV-1, it has been hypothesized that sera from convalescent SARS-CoV-1 patients might show some efficacy in cross-neutralizing SARS-CoV-2-S-driven entry [70].
However, despite the high level of sequence homology, certain protein structures might be immunologically distinct, which would be likely to prohibit effective cross-neutralization across different SARS species [172].
Consequently, proteomic analyses of SARS-CoV-1 might also provide some essential information regarding the new pathogen [173,174].
Considering the paucity of omics-level big data sets for SARS-CoV-2 currently available, existing data hubs that contain information for other coronaviruses such as UniProt [175], NCBI Genome Database [176], The Immune Epitope Database and Analysis Resource [177], and The Virus Pathogen Resource [178] will serve as useful resources for comparative bioinformatics research of SARS-CoV-2.
Using such databases, the systems-level reconstruction of protein-protein interaction networks will enable the generation of hypotheses about the mechanism of action of SARS-CoV-2 and suggest potential drug targets.
In an initial study [179], 26 of the 29 SARS-CoV-2 proteins were cloned and expressed in HEK293T kidney cells, allowing for the identification of 332 high-confidence human proteins interacting with them.
Notably, this study suggested that SARS-CoV-2 interacts with innate immunity pathways.
Ranking pathogens by the similarity between their interactomes and that of SARS-CoV-2 suggested West Nile virus, Mycobacterium tuberculosis, and human papillomavirus infections as the top three hits.
Therefore, given the lung symptoms associated with COVID-19, the Mycobacterium tuberculosis host-pathogen interactome in particular might provide new insights to the mechanism of SARS-CoV-2 infection.
Additionally, it was suggested that the envelope protein, E, could disrupt host bromodomain-containing proteins, i.e., BRD2 and BRD4, that bind to histones, and the spike protein could likely intervene in viral fusion by modulating the GOLGA7-ZDHHC5 acyl-transferase complex to increase palmitoylation, which is a post-translational modification that affects how proteins interact with membranes [180].
Another study [181] used patient-derived peripheral blood mononuclear cells to identify 251 host proteins targeted by SARS-CoV-2.
This study also reported that more than 200 host proteins were disrupted following infection.
In particular, a network analysis showed that nsp9 and nsp10 interacted with NF-Kappa-B-Repressing Factor, which encodes a transcriptional repressor that mediates repression of genes responsive to Nuclear Factor kappa-light-chain-enhancer of activated B-cells.
These genes are important to pro-, and potentially also anti-, inflammatory signaling [182].
This finding could explain the exacerbation of the immune response that shapes the pathology and the high cytokine levels characteristic of COVID-19, possibly due to the chemotaxis of neutrophils mediated by IL-8 and IL-6.
Finally, it was suggested [183] that the E protein of both SARS-CoV-1 and SARS-CoV-2 has a conserved Bcl-2 Homology 3-like motif, which could inhibit anti-apoptosis proteins, e.g., BCL2, and trigger the apoptosis of T cells.
Several compounds are known to disrupt the host-pathogen protein interactome, largely through the inhibition of host proteins.
Therefore, this research identifies candidate targets for intervention and suggests that drugs modulating protein-level interactions between virus and host could be relevant to treating COVID-19.
By revealing which genes are perturbed during SARS-CoV-2 infection, proteomics-based analyses can thus provide novel insights into host-virus interaction and serve to generate new avenues of investigation for therapeutics.
2.7 Viral Virulence
Like that of SARS-CoV-1, the entry of SARS-CoV-2 into host cells is mediated by interactions between the viral spike glycoprotein, S, and human ACE2 (hACE2) [67,70,184,185,186,187,188,189].
Differences in how the S proteins of the two viruses interact with hACE2 could partially account for the increased transmissibility of SARS-CoV-2.
Recent studies have reported conflicting binding constants for the S-hACE2 interaction, though they have agreed that the SARS-CoV-2 S protein binds with equal, if not greater, affinity than the SARS-CoV-1 S protein does [54,67,187].
The C-terminal domain of the SARS-CoV-2 S protein in particular was identified as the key region of the virus that interacts with hACE2, and the crystal structure of the C-terminal domain of the SARS-CoV-2 S protein in complex with hACE2 reveals stronger interaction and a higher affinity for receptor binding than that of SARS-CoV-1 [188].
Among the 14 key binding residues identified in the SARS-CoV-1 S protein, eight are conserved in SARS-CoV-2, and the remaining six are semi-conservatively substituted, potentially explaining variation in binding affinity [67,187].
Recent crystal structures have shown that the receptor binding domain (RBD) of the SARS-CoV-2 S protein, like that of other coronaviruses, undergoes stochastic hinge-like movement that flips it from a “closed” conformation, in which key binding residues are hidden at the interface between protomers, to an “open” one [54,67].
Because the RBD plays such a critical role in viral entry, blocking its interaction with ACE2 could represent a promising therapeutic approach.
Nevertheless, despite the high structural homology between the SARS-CoV-2 RBD and that of SARS-CoV-1, monoclonal antibodies targeting SARS-CoV-1 RBD failed to bind to SARS-CoV-2-RBD [54].
However, in early research, sera from convalescent SARS patients were found to inhibit SARS-CoV-2 viral entry in vitro, albeit with lower efficiency than it inhibited SARS-CoV-1 [70].
Comparative genomic analysis reveals that several regions of the coronavirus genome are likely critical to virulence.
The S1 domain of the spike protein, which contains the receptor binding motif, evolves more rapidly than S’s S2 domain [60,61].
However, even within the S1 domain, some regions are more conserved than others, with the receptors in S1’s N-terminal domain (S1-NTD) evolving more rapidly than those in its C-terminal domain (S1-CTD) [61].
Both S1-NTD and S1-CTD are involved in receptor binding and can function as RBDs to bind proteins and sugars [60], but RBDs in the S1-NTD typically bind to sugars, while those in the S1-CTD recognize protein receptors [50].
Viral receptors show higher affinity with protein receptors than sugar receptors [50], which suggests that positive selection on or relaxed conservation of the S1-NTD might reduce the risk of a deleterious mutation that would prevent binding.
The SARS-CoV-2 S protein also contains an RRAR furin recognition site at the S1/S2 junction [54,67], setting it apart from both bat coronavirus RaTG13, with which it shares 96% genome sequence identity, and SARS-CoV-1 [15].
Such furin cleavage sites are commonly found in highly virulent influenza viruses, and as such may contribute to the heightened pathogenicity of SARS-CoV-2 [190,191].
The ongoing evolution of the spike protein can be seen from the genomic data.
For example, the mutation D614G became dominant by the end of May 2020, soon after its initial appearance in mid-March [192,193], and a variant carrying two mutations (N501Y and 69–70del) that was first observed in the UK in October 2020 [194] has quickly spread around the world [195,196].
Variants may differ in transmissibility [197,198].
Effective cell entry is a critical component to pathogenesis and therefore an important process to understand when examining possible therapeutics.
2.8 Mechanism of Transmission
Once a human host is infected with a virus, person-to-person viral transmission can occur through several possible mechanisms.
The primary mechanisms associated with respiratory viruses are contact, droplet, and aerosol transmission [199].
Contact transmission can occur through either contact with a contagious person or contact with active viral particles on a contaminated surface [200].
This latter mode of transmission is also called fomite transmission [201].
Viral particles can enter the body if they then come in contact with the oral, nasal, eye, or other mucus membranes [200].
Droplet transmission occurs when a contagious individual sneezes, coughs, or exhales and produces respiratory droplets that can contain a large number of viral particles [200].
Contact with these droplets can occur through direct exposure to the droplets, such as breathing in droplets produced by a sneeze [200].
The droplets can also potentially settle on a surface and contribute to fomite transmission [200].
Aerosol transmission refers to much smaller particles (less than 5 micrometers) that are also produced by sneezing, coughing, or exhaling [199,200].
The small size of these particles allows them to remain suspended over a longer period of time and potentially to be moved by air currents [200].
Additionally, viral particles deposited on surfaces via large respiratory droplets can also later be aerosolized [200].
Droplet and/or contact transmission are both well-accepted modes of transmission for many viruses associated with common human illnesses, including influenza and rhinovirus [200].
The extent to which aerosol transmission contributes to the spread of respiratory viruses is less clear.
In influenza A, for example, viral particles can be detected in aerosols produced by infected individuals, but the extent to which these particles drive the spread of influenza A infection remains under debate [199,200,202,203,204].
Regardless of its role in the spread of influenza A, however, aerosol transmission likely played a role in outbreaks such as the 1918 Spanish Influenza (H1N1) and 2009 “swine flu” (pH1N1) [204].
Contact, droplet, and aerosol transmission are therefore all worth evaluating when considering possible modes of transmission for a respiratory virus like SARS-CoV-2.
All three of these mechanisms have been identified as possible contributors to the transmission of HCoVs [200], including the highly pathogenic coronaviruses SARS-CoV-1 and MERS-CoV [31,205].
Transmission of SARS-CoV-1 is thought to proceed primarily through droplet transmission, but aerosol transmission is also considered possible [200], and fomite transmission may have also played an important role in some outbreaks [206].
Similarly, the primary mechanism of MERS transmission is thought to be droplets because inter-individual transmission appears to be associated with close interpersonal contact (e.g., household or healthcare settings), but aerosolized particles of the MERS virus have been reported to persist much more robustly than influenza A under a range of environmental conditions [207,208].
While droplet-based and contact transmission were initially put forward as the greatest concern for the spread of SARS-CoV-2 [209], as additional information has emerged, the possibility of aerosol transmission has also been raised [210,211,212].
For example, the detection of SARS-CoV-2 viral particles in air samples taken from hospitals treating COVID-19 patients led to the concern that the virus could be spreading via aerosols [213].
The stability of the virus both in aerosols and on a variety of surfaces appeared similar to that of SARS-CoV-1 [211].
However, while the possibility of aerosol transmission seems plausible, the evidence suggests that droplet transmission is the dominant mechanism driving the spread of the virus [214], and the risk of fomite transmission under real-world conditions is likely to be substantially lower than the conditions used for experimental analyses [215].
These mechanisms may differ in their relevance to different types of transmission events, such as transmission within households, nosocomial transmissions, and transmission in indoor versus outdoor spaces.
2.8.1 Symptoms and Viral Spread
Other aspects of pathogenesis are also important to understanding how the virus spreads, especially the relationship between symptoms, viral shedding, and contagiousness.
Symptoms associated with reported cases of COVID-19 range from mild to severe [47], but some individuals who contract COVID-19 remain asymptomatic throughout the duration of the illness [216].
The incubation period, or the time period between exposure and the onset of symptoms, has been estimated at five to eight days, with means of 4.91 (95% confidence interval (CI) 4.35-5.69) and 7.54 (95% CI 6.76-8.56) reported in two different Asian cities and a median of 5 (IQR 1 to 6) reported in a small number of patients in a Beijing hospital [217,218].
However, the exact relationship between contagiousness and viral shedding remains unclear.
Estimates suggest that viral shedding can, in some cases, begin as early as 12.3 days (95% CI 5.9-17.0) before the onset of symptoms, although this was found to be very rare, with less than 0.1% of transmission events occurring 7 or more days before symptom onset [219].
Transmissibility appeared to peak around the onset of symptoms (95% CI -0.9 - 0.9 days), and only 44% (95% CI 30–57%) of transmission events were estimated to occur from presymptomatic contacts [219].
As these trends became apparent, concerns arose due to the potential for individuals who did not yet show symptoms to transmit the virus [220].
Recovered individuals may also be able to transmit the virus after their symptoms cease.
Estimates of the communicable period based on twenty-four individuals who tested positive for SARS-CoV-2 prior to or without developing symptoms estimated that individuals may be contagious for one to twenty-one days, but they note that this estimate may be low [216].
In an early study, viral nucleic acids were reported to remain at observable levels in the respiratory specimens of recovering hospitalized COVID-19 patients for a median of 20 days and with a maximum observed duration through 37 days, when data collection for the study ceased [38].
As more estimates of the duration of viral shedding are released, they are beginning to converge around approximately three weeks from first positive PCR test and/or onset of symptoms (which, if present, are usually identified within three days of the initial PCR test).
For example, in later studies, viral shedding was reported for up to 28 days following symptom onset [221] and for one to 24 days from first positive PCR test, with a median of 12 days [98].
On the other hand, almost 70% of patients were reported to still have symptoms at the time that viral shedding ceased, although all symptoms reduced in prevalence between onset and cessation of viral shedding [222].
The median time that elapsed between the onset of symptoms and cessation of viral RNA shedding was 23 days and between first positive PCR test and cessation of viral shedding was 17 days [222].
The fact that this study reported symptom onset to predate the first positive PCR test by an average of three days, however, suggests that there may be some methodological differences between it and related studies.
Furthermore, an analysis of residents of a nursing home with a known SARS-CoV-2 case measured similar viral load in residents who were asymptomatic regardless of whether they later developed symptoms, and the load in the asymptomatic residents was comparable to that of residents who displayed either typical of atypical symptoms [223].
Taken together, these results suggest that the presence or absence of symptoms are not reliable predictors of viral shedding or of SARS-CoV-2 status (e.g, [224]).
However, it should be noted that viral shedding is not necessarily a robust indicator of contagiousness.
The risk of spreading the infection was low after ten days from the onset of symptoms, as viral load in sputum was found to be unlikely to pose a significant risk based on efforts to culture samples in vitro[221].
The relationship between symptoms, detectable levels of the virus, and risk of viral spread is therefore complex.
The extent to which asymptomatic or presymptomatic individuals are able to transmit SARS-CoV-2 has been a question of high scientific and community interest.
Early reports (February and March 2020) described transmission from presymptomatic SARS-CoV-2-positive individuals to close family contacts [225,226].
One of these reports [226] also included a description of an individual who tested positive for SARS-CoV-2 but never developed symptoms.
Later analyses also sought to estimate the proportion of infections that could be traced back to a presymptomatic or asymptomatic individual (e.g., [227]).
Estimates of the proportion of individuals with asymptomatic infections have varied widely.
The proportion of asymptomatic individuals on board the Diamond Princess cruise ship, which was the site of an early COVID-19 outbreak, was estimated at 17.9% [228].
In contrast, a model using the prevalence of antibodies among residents of Wuhan, China estimated a much higher rate of asymptomatic cases, at approximately 7 in 8, or 87.5% [229].
An analysis of the populations of care homes in London found that, among the residents (median age 85), the rate of asymptomatic infection was 43.8%, and among the caretakers (median age 47), the rate was 49.1% [230].
The duration of viral shedding may also be longer in individuals with asymptomatic cases of COVID-19 compared to those who do show symptoms [231].
As a result, the potential for individuals who do not know they have COVID-19 to spread the virus raises significant concerns.
In Singapore and Tianjin, two cities studied to estimate incubation period, an estimated 40-50% and 60-80% of cases, respectively, were considered to be caused by contact with asymptomatic individuals [217].
An analysis of viral spread in the Italian town of Vo’, which was the site of an early COVID-19 outbreak, revealed that 42.5% of cases were asymptomatic and that the rate was similar across age groups [232].
The argument was thus made that the town’s lockdown was imperative for controlling the spread of COVID-19 because it isolated asymptomatic individuals.
While more models are likely to emerge to better explore the effect of asymptomatic individuals on SARS-CoV-2 transmission, these results suggest that strategies for identifying and containing asymptomatic but contagious individuals are important for managing community spread.
2.8.2 Estimating the Fatality Rate
Estimating the occurrence of asymptomatic and mild COVID-19 cases is important to identifying the mortality rate associated with COVID-19.
The mortality rate of greatest interest would be the total number of fatalities as a fraction of the total number of people infected.
One commonly reported metric is the case fatality rate (CFR), which compares the number of COVID-19 related deaths to the number of confirmed or suspected cases.
However, in locations without universal testing protocols, it is impossible to identify all infected individuals because so many asymptomatic or mild cases go undetected.
Therefore, a more informative metric is the infection fatality rate (IFR), which compares the known deaths to the estimated number of cases.
It thus requires the same numerator as CFR, but divides by an approximation of the total number of cases rather than only the observed/suspected cases.
IFR varies regionally, with some locations observed to have IFRs as low as 0.17% while others are as high as 1.7% [233].
Estimates of CFR at the national and continental level and IFR at the content level is maintained by the Centre for Evidence-Based Medicine [234].
Several meta-analyses have also sought to estimate IFR at the global scale.
These estimates have varied; one peer-reviewed study aggregated data from 24 other studies and estimated IFR at 0.68% (95% CI 0.53%–0.82%), but a preprint that aggregated data from 139 countries calculated a global IFR of 1.04% (95% CI 0.77%-1.38%) when false negatives were considered in the model [233,235].
A similar prevalence estimate was identified through a repeated cross-sectional serosurvey conducted in New York City that estimated the IFR as 0.97% [236].
Examination of serosurvey-based estimates of IFR identified convergence on a global IFR estimate of 0.60% (95% CI 0.42%–0.77%) [233].
All of these studies note that IFR varies widely by location, and it is also expected to vary with demographic and health-related variables such as age, sex, prevalence of comorbidities, and access to healthcare and testing [237].
Estimates of infection rates are becoming more feasible as more data becomes available for modeling and will be bolstered as serological testing becomes more common and more widely available.
2.9 Dynamics of Transmission
Disease spread dynamics can be estimated using R0, the basic reproduction number, and Rt, the effective reproduction number.
Accurate estimates of both are crucial to understanding the dynamics of infection and to predicting the effects of different interventions.
R0 is the average number of new (secondary) infections caused by one infected person, assuming a wholly susceptible population [238] and is one of the most important epidemiological parameters [239].
A simple mechanistic model used to describe infectious disease dynamics is a susceptible-infected-recovered compartmental model [240,241].
In this model, individuals move through three states: susceptible, infected, and recovered; two parameters, \(\gamma\) and \(\beta\), specify the rate at which the infectious recover, and the infection transmission rate, respectively, and R0 is estimated as the ratio of \(\beta\) and \(\gamma\)[239,242].
A pathogen can invade a susceptible population only if R0 > 1 [239,243].
The spread of an infectious disease at a particular time t can be quantified by Rt, the effective reproduction number, which assumes that part of the population has already recovered (and thus gained immunity to reinfection) or that mitigating interventions have been put into place.
For example, if only a fraction St of the population is still susceptible, Rt = St x R0.
When Rt is greater than 1, an epidemic grows (i.e., the proportion of the population that is infectious increases); when Rt is less than 1, the proportion of the population that is infectious decreases.
R0 and Rt can be estimated directly from epidemiological data or inferred using susceptible-infected-recovered-type models.
To accurately capture the dynamics of SARS-CoV-2, the addition of a fourth compartment, i.e. a susceptible-exposed-infectious-recovered model may be appropriate.
Estimates of R0 for COVID-19 lie in the range R0=1.4-6.5 [244,245,246].
Variation in R0 is expected between different populations, and the estimated values of R0 discussed below are for specific populations in specific environments.
The different estimates of R0 should not necessarily be interpreted as a range of estimates of the same underlying parameter.
In one study of international cases, the predicted value was R0=1.7 [247].
In China (both Hubei province and nationwide), the value was predicted to lie in the range R0=2.0-3.6 [244,248,249].
Another estimate based on a cruise ship where an outbreak occurred predicted R0=2.28 [250].
Susceptible-exposed-infectious-recovered model-derived estimates of R0 range from 2.0 - 6.5 in China [251,252,253,254] to R0=4.8 in France [255].
Using the same model as for the French population, a study estimated R0=2.6 in South Korea [255], which is consistent with other studies [256].
From a meta-analysis of studies estimating R0, [245] the median R0 was estimated to be 2.79 (IQR 1.16) based on twelve studies published between January 1 and February 7, 2020.
Inference of the effective reproduction number can provide insight into how populations respond to an infection and the effectiveness of interventions.
In China, Rt was predicted to lie in the range 1.6-2.6 in January 2020, before travel restrictions [257].
Rt decreased from 2.35 one week before travel restrictions were imposed (January 23, 2020), to 1.05 one week after.
Using their model, the authors also estimated the probability of new outbreaks occurring.
Assuming individual-level variation in transmission comparable to that of MERS or SARS, the probability of a single individual exporting the virus and causing a large outbreak is 17-25%, and assuming variation like that of SARS and transmission patterns like those observed for COVID-19 in Wuhan, the probability of a large outbreak occurring after ≥4 infections exist at a new location is greater than 50%.
An independent study came to similar conclusions, finding Rt=2.38 in the two-week period before January 23 with a decrease to Rt = 1.34 (using data from January 24 to February 3) or Rt=0.98 (using data from January 24 to February 8) [246].
In South Korea, Rt was inferred for February through March 2020 in two cities, Daegu (the center of the outbreak) and Seoul [256].
Metro data was also analyzed to estimate the effects of social distancing measures.
Rt decreased in Daegu from around 3 to <1 over the period that social distancing measures were introduced.
In Seoul, Rt decreased slightly, but remained close to 1 (and larger than Rt in Daegu).
These findings indicate that social distancing measures appeared to be effective in containing the infection in Daegu, but in Seoul, Rt remained above 1, meaning secondary outbreaks remained possible.
The study also shows the importance of region-specific analysis: the large decline in case load nationwide was mainly due to the Daegu region and could mask persistence of the epidemic in other regions, such as Seoul and Gyeonggi-do.
In Iran, estimates of Rt declined from 4.86 in the first week to 2.1 by the fourth week after the first cases were reported [258].
In Europe, analysis of 11 countries inferred the dynamics of Rt over a time range from the beginning of the outbreak until March 28, 2020, by which point most countries had implemented major interventions (such as stay-at-home orders, public gathering bans, and school closures) [259].
Across all countries, the mean Rt before interventions began was estimated as 3.87; Rt varied considerably, from below 3 in Norway to above 4.5 in Spain.
After interventions, Rt decreased by an average of 64% across all countries, with mean Rt=1.43.
The lowest predicted value was 0.97 for Norway and the highest was 2.64 for Sweden, which could be related to the fact that Sweden did not implement social distancing measures on the same scale as other countries.
The study concludes that while large changes in Rt are observed, it is too early to tell whether the interventions put into place are sufficient to decrease Rt below 1.
More generally, population-level epidemic dynamics can be both observed and modeled [242].
Data and empirically determined biological mechanisms inform models, while models can be used to try to understand data and systems of interest or to make predictions about possible future dynamics, such as the estimation of capacity needs [260] or the comparison of predicted outcomes among prevention and control strategies [261,262].
Many current efforts to model Rt have also led to tools that assist the visualization of estimates in real time or over recent intervals [263,264].
These are valuable resources, yet it is also important to note that the estimates arise from models containing many assumptions and are dependent on the quality of the data they use, which varies widely by region.
2.10 Molecular Signatures, Transmission, and Variants of Concern
Genetic variation in SARS-CoV-2 has been used to elucidate patterns over time and space.
Mutations observed in individual SARS-CoV-2 genome sequences can be used to trace transmission patterns and have provided insights during outbreak investigations [17,265,266].
Similar mutations observed in several patients may indicate that the patients belong to the same transmission group.
The tracking of SARS-CoV-2 mutations is recognized as an essential tool for controlling future outbreaks and tracing the path of the spread of SARS-CoV-2.
Efforts vary widely by country: the UK has coordinated a national database of viral genomes [267]; no such coordination has been achieved in the USA.
Several studies used phylogenetic analysis to determine the source of local COVID-19 outbreaks in Connecticut (USA), [268], the New York City area (USA) [269], and Iceland [270].
There is an ongoing effort to collect SARS-CoV-2 genomes throughout the COVID-19 outbreak, and as of January 18, 2021 more than 381,000 genome sequences have been collected from patients.
The sequencing data can be found at GISAID [271], NCBI [272], and COVID-19 data portal [273].
Some SARS-CoV-2 variants, which we review in more detail separately [???], may be associated with increased transmission of SARS-CoV-2.
For example, coordinated sequencing efforts of SARS-CoV-2 genomes in the U.K. were instrumental in detecting B.1.1.7/VOC 202012/01, a variant of concern (VOC) that, with an estimated R0 of 1.4, is associated with approximately 50% increased transmission due to genetic mutations in the Spike gene that modify the Spike protein [274,275,276].
This variant has now spread to at least 93 other countries [277], and other studies investigating its transmission have occurred.
In the U.S. between December 2020 and January 2021, it was estimated that B.1.1.7 had an increased transmission of 35-45% relative to common SARS-CoV-2 variants at the time, and B.1.1.7 is now on its way to becoming the dominant SARS-CoV-2 variant in the U.S. [278].
Notably, the latest analyses in the U.K. reports that there may also be an increased risk of disease severity associated with B.1.1.7 [279] and potentially a 35% increased risk of death [280] compared to other common variants of SARS-CoV-2.
However, further studies are required to confirm these associations.
Other variants meriting further investigation have also been identified worldwide, including the P.1 lineage associated with outbreaks in Brazil and the B.1.351 variant first identified in South Africa.
Currently, it is not known whether these lineages have increased transmissibility.
However, the P.1. lineage and the B.1.351 variant do share similar independently acquired mutations with B.1.1.7 such as N501Y [281,282].
Indeed, these novel variants are being investigated for potential resistance to vaccines and antibody treatments such as Bamlanivimab, but to date it seems that these agents are still effective, albeit with reduced efficacy in the case of the Pfizer-BioNTech vaccine [283].
As a consequence of reliance on targeting the SARS-CoV-2 Spike protein for many therapeutic and prophylactic strategies, increased surveillance is required to rapidly identify and prevent the spread of novel SARS-CoV-2 variants with alterations to the Spike protein.
2.11 Conclusions
The novel coronavirus SARS-CoV-2 is the third HCoV to emerge in the 21st century, and research into previous HCoVs has provided a strong foundation for characterizing the pathogenesis and transmission of SARS-CoV-2.
Critical insights into how the virus interacts with human cells have been gained from previous research into HCoVs and other viral infections.
As with other HCoVs, the immune response to SARS-CoV-2 is likely driven by detection of its spike protein, which allows it to enter cells through ACE2.
Epithelial cells have also emerged as the major cellular target of the virus, contextualizing the respiratory and gastrointestinal symptoms that are frequently observed in COVID-19.
Many of the mechanisms that facilitate the pathogenesis of SARS-CoV-2 are currently under consideration as possible targets for the treatment or prevention of COVID-19.
Research in other viruses also provides a foundation for understanding the transmission of SARS-CoV-2 among people and can therefore inform efforts to control the virus’s spread.
The extent to which aerosol and fomite transmission contribute to the spread of SARS-CoV-2 remains a question: in general, much like SARS-CoV-1 and MERS-CoV, this virus seems to be spread primarily by droplet transmission.
Asymptomatic transmission was also a concern in the SARS outbreak of 2002-03 and, as in current pandemic, presented challenges for estimating rates of infection [284].
However, in the current pandemic, we have been fortunate to be able to build on top of 18 years of SARS-CoV-1 research in order to rapidly ascertain the identity and behavior of the virus.
Even with the background obtained from research in SARS and MERS, COVID-19 has revealed itself to be a complex and difficult-to-characterize disease that has many possible presentations that vary with age.
Variability in presentation, including cases with no respiratory symptoms or with no symptoms altogether, were also reported during the SARS epidemic at the beginning of the 21st century [284].
The variability of both which symptoms present and their severity have presented challenges for public health agencies seeking to provide clear recommendations regarding which symptoms indicate SARS-CoV-2 infection and should prompt isolation.
Asymptomatic cases add complexity both to efforts to estimate statistics such as R0 and Rt, which are critical to understanding the transmission of the virus, and IFR, which is an important component of understanding its impact on a given population.
The development of diagnostic technologies over the course of the pandemic has facilitated more accurate identification, including of asymptomatic cases.
As more cases have been diagnosed, the health conditions and patient characteristics associated with more severe infection have also become more clear, although there are likely to be significant sociocultural elements that also influence these outcomes.
While many efforts have focused on adults, and especially older adults because of the susceptibility of this demographic, additional research is needed to understand the presentation of COVID-19 and MIS-C in pediatric patients.
As more information is uncovered about the pathogenesis of HCoV and SARS-CoV-2 specifically, the diverse symptomatology of COVID-19 has and likely will continue to conform with the ever-broadening understanding of how SARS-CoV-2 functions within a human host.
While the SARS-CoV-2 virus is very similar to other HCoV in several ways, including in its genomic structure and the structure of the virus itself, there are also some differences that may account for differences in the COVID-19 pandemic compared to the SARS and MERS epidemics of the past two decades.
The R0 of SARS-CoV-2 has been estimated to be similar to SARS-CoV-1 but much higher than that of MERS-CoV [33,33].
While the structures of the viruses are very similar, evolution among these species may account for differences in their transmissibility and virulence.
For example, the acquisition of a furin cleavage site the S1/S2 boundary within the SARS-CoV-2 S protein may be associated with increased virulence.
Additionally, concerns have been raised about the accumulation of mutations within the SARS-CoV-2 species itself, and whether these could influence virulence.
The coming of age of genomic technologies has made these types of analyses feasible, and genomics research characterizing changes in SARS-CoV-2 along with temporal and spatial movement is likely to provide additional insights into whether within-species evolution influences the effect of the virus on the human host.
Additionally, the rapid development of sequencing technologies over the past decade has made it possible to rapidly characterize the host response to the virus.
For example, proteomics analysis of patient-derived cells revealed candidate genes whose regulation is altered by SARS-CoV-2 infection, suggesting possible approaches for pharmaceutical invention and providing insight into which systems are likely to be disrupted in COVID-19 [181].
As more patient data becomes available, the biotechnological advances of the 2000s are expected to allow for more rapid identification of potential drug targets than was feasible during the SARS, or even MERS, pandemic.
Thus, though the COVID-19 crisis is still evolving, the insights acquired over the past 20 years of HCoV research have provided a solid foundation for understanding the SARS-CoV-2 virus and the disease it causes.
As the scientific community continues to respond to COVID-19 and to elucidate more of the relationships between viral pathogenesis, transmission, and symptomatology, and as more data about the regulatory shifts associated with COVID-19 become available, this understanding will no doubt continue to evolve and to reveal additional connections among virology, pathogenesis, and health.
As additional information becomes available, this review will be updated to reflect the changing state of research in this area.
At present, understanding the SARS-CoV-2 virus and its pathogenesis is critical to a holistic understanding of the COVID-19 pandemic.
3 Evolutionary and Genomic Analysis of SARS-CoV-2
3.1 Abstract
3.2 Introduction
3.3 Initial Characterization of SARS-CoV-2
The first genome sequence of the virus was released on January 3, 2020.
It revealed that the cluster of pneumonia cases seen in Wuhan were caused by a novel coronavirus [13].
Multiple research groups have drafted the genome sequence of SARS-CoV-2 based on sequences developed from clinical samples collected from the lower respiratory tract, namely bronchoalveolar lavage fluid (BALF), and the upper respiratory tract, in the form of throat and nasopharyngeal swabs [14,15,16].
Analysis of the SARS-CoV-2 genome revealed significant sequence homology with two coronaviruses known to infect humans, with about 79% identity to SARS-CoV-1 and 50% to MERS-CoV [16].
However, in this analysis, the highest degree of similarity was observed between SARS-CoV-2 and bat-derived SARS-like coronaviruses (bat-SL-CoVZC45 and bat-SL-CoVZXC21) [16].
Other analyses have reported even greater similarity between SARS-CoV-2 and the bat coronavirus BatCoV-RaTG13, with identity as high as 96.2% [15,17], and the closely related pangolin coronavirus [18].
This evidence therefore suggests the SARS-CoV-2 virus is the result of zoonotic transfer of a virus from bats to humans.
Nevertheless, some fragments between SARS-CoV-2 and RATG13 differ by up to 17%, suggesting a complex natural selection process during zoonotic transfer.
While the S region is highly similar to that of viruses found in pangolins [18], there is no consensus about the origin of S in SARS-CoV-2, as it could potentially be the result either of recombination or coevolution [17,19].
Though the intermediate host serving as the source for the zoonotic introduction of SARS-CoV-2 to human populations has not yet been identified, the SARS-CoV-2 virus has been placed within the coronavirus phylogeny through comparative genomic analyses.
Genomic analyses and comparisons to other known coronaviruses suggest that SARS-CoV-2 is unlikely to have originated in a laboratory – either purposely engineered and released, or escaped – and instead evolved naturally in an animal host [20].
Indeed, the World Health Organization (WHO) have published their intentions to thoroughly investigate the origins of SARS-CoV-2 [285].
While the position of the SARS-CoV-2 virus within the coronavirus phylogeny has been largely resolved, the functional consequences of molecular variation between this virus and other viruses, such as its bat and pangolin sister taxa or SARS-CoV-1, remain unknown [20].
Fortunately, the basic genome structure of coronaviruses is highly conserved, and insight into the mechanisms the virus uses to enter cells, replicate, and spread is available from prior research on coronaviruses, which has been instrumental in the mobilization of global research to understand the biology of SARS-CoV-2.
Additionally, worldwide sequencing of viral samples has provided some preliminary insights into possible mechanisms of adaptation in the virus and the detection of novel variants, and omics-based analysis of patient samples has elucidated some of the biological changes the virus induces in its human hosts.
3.4 Coronaviruses and Animal Hosts
Coronaviruses have long been known to infect animals and have been the subject of veterinary medical investigations and vaccine development efforts due to their effect on the health of companion and agricultural animals [21].
Most coronaviruses show little to no transmission in humans.
However, it is thought that approximately one-third of common cold infections are caused by four seasonal human coronaviruses (HCoV): Human coronavirus 229E (HCoV-229E), Human coronavirus NL63 (HCoV-NL63), Human coronavirus OC43 (HCoV-OC43), and Human coronavirus HKU1 (HCoV-HKU1) [22,23,286].
The first HCoV were identified in the 1960s: HCoV-229E in 1965 [24] and HCoV-OC43 in 1967 [25].
Both of these viruses typically cause cold-like symptoms, including upper and lower respiratory infections [26,27,287], but they have also been associated with gastrointestinal symptoms [288].
Two additional HCoV were subsequently identified [28,29].
In 2003, HCoV-NL63 [28] was first identified in a 7-month-old infant and then in clinical specimens collected from seven additional patients, five of whom were infants younger than 1 year old and the remainder of whom were adults.
CoV-HKU1 was identified in samples collected from a 71-year-old pneumonia patient in 2004 and then found in samples collected from a second adult patient [29].
These viruses are associated with respiratory diseases of varying severity, ranging from common cold to severe pneumonia, with severe symptoms mostly observed in immunocompromised individuals [30], and also have gastrointestinal involvement in some cases [288].
In addition to these relatively mild HCoV, however, highly pathogenic human coronaviruses have been identified, including Severe acute respiratory syndrome-related coronavirus (SARS-CoV or SARS-CoV-1) and Middle East respiratory syndrome-related coronavirus (MERS-CoV) [22,31,32].
At the time that SARS-CoV-1 emerged in the early 2000s, no HCoV had been identified in almost 40 years [31].
The first case of SARS was reported in November 2002 in the Guangdong Province of China, and over the following month, the disease spread more widely within China and then into several countries across multiple continents [31,33].
Unlike previously identified HCoV, SARS was much more severe, with an estimated death rate of 9.5% [33].
It was also highly contagious via droplet transmission, with a basic reproduction number (R0) of 4 (i.e., each person infected was estimated to infect four other people) [33].
However, the identity of the virus behind the infection remained unknown until April of 2003, when the SARS-CoV-1 virus was identified through a worldwide scientific effort spearheaded by the WHO [31].
SARS-CoV-1 belonged to a distinct lineage from the two other HCoV known at the time [33].
By July 2003, the SARS outbreak was officially determined to be under control, with the success credited to infection management practices [31].
A decade later, a second outbreak of severe respiratory illness associated with a coronavirus emerged, this time in the Arabian Peninsula.
This disease, known as Middle East respiratory syndrome (MERS), was linked to another novel coronavirus, MERS-CoV.
The fatality rate associated with MERS is much higher than that of SARS, at almost 35%, but the disease is much less easily transmitted, with an R0 of 1 [33].
Although MERS is still circulating, its low reproduction number has allowed for its spread to be contained [33].
The COVID-19 pandemic is thus associated with the seventh HCoV to be identified and the fifth since the turn of the millennium, though additional HCoVs may be in circulation but remain undetected.
SARS-CoV-1 and MERS-CoV were ultimately managed largely through infection management practices (e.g., mask wearing) and properties of the virus itself (i.e., low rate of transmission), respectively [31,33].
Vaccines were not used to control either outbreak, although vaccine development programs were established for SARS-CoV-1 [34].
In general, care for SARS and MERS patients focuses on supportive care and symptom management [33].
Clinical treatments for SARS and MERS developed during the outbreaks generally do not have strong evidence supporting their use.
Common treatments included Ribavirin, an antiviral, often in combination with corticosteroids or sometimes interferon (IFN) medications, which would both be expected to have immunomodulatory effects [31].
However, retrospective and in vitro analyses have reported inconclusive results of these treatments on SARS and the SARS-CoV-1 virus, respectively [31].
IFNs and Ribavirin have shown promise in in vitro analyses of MERS, but their clinical effectiveness remains unknown [31].
Therefore, only limited strategies can be adopted for the pharmaceutical management of COVID-19 from previous severe HCoV infections.
Research in response to prior outbreaks of HCoV-borne infections, such as SARS and MERS, have, however, provided a strong foundation for hypotheses about the pathogenesis of SARS-CoV-2 as well as potential diagnostic and therapeutic approaches.
3.5 Zoonotic Transfer of Coronaviruses
3.6 Evolution of the SARS-CoV-2 Virus
3.6.1 Emergence of SARS-CoV-2
3.6.2 Evolution of SARS-CoV-2 Variants
Evolution in SARS-CoV-2 has also been observed over a short timescale.
After zoonotic transfer, SARS-CoV-2 continued evolving in the human population [265].
The SARS-CoV-2 mutation rate is moderate compared to other RNA viruses [266], which likely restricts the pace of evolution in SARS-CoV-2.
Nevertheless, genomic analyses have yielded statistical evidence of ongoing evolution.
Initially, two known variants of the spike protein emerged that differed by a single amino acid at position 614 (G614 and D614), and there is evidence that G614 had become more prevalent than D614 by June 2020 [192].
While there is a hypothesis that this genomic change increased the SARS-CoV-2 infectivity and virulence, this hypothesis has not yet been tested due to a lack of data [289].
Another study [266] identified 198 recurrent mutations in a dataset of 7,666 curated sequences.
This pattern of convergent evolution at some sites could indicate that certain mutations confer an adaptive advantage.
While it is evident that SARS-CoV-2 exhibits moderate potential for ongoing and future evolution, the relationship between mutations and pathogenicity is not yet known.
Additional data is needed in order to understand patterns of evolutionary change and whether they are likely to affect virulence.
Several factors could promote the evolution of SARS-CoV-2, including host immunodeficiency and transient exposure to antibodies directed against SARS-CoV-2 proteins.
A single case study of SARS-CoV-2 infection in an immunocompromised female with chronic lymphocytic leukemia and hypogammaglobulinemia [290] suggested that an accelerated evolution of the virus could occur in conditions of immunodeficiency.
A first administration of convalescent plasma did not clear the virus, and an ensuing increase in the genomic diversity in the samples was observed, suggesting an accelerated evolution due to selection pressure.
A second administration of convalescent plasma cleared the virus from the host 105 days after the initial diagnosis.
However, throughout the duration of infection, the patient was asymptomatic but contagious.
A second single case study in a 45-year old male with antiphospholipid syndrome [291] confirmed the earlier results, providing evidence of persistent COVID-19 symptoms in an immunocompromised patient for 154 days following diagnosis, ultimately leading to the death of patient.
The treatments administered included remdesivir and the Regeneron anti-spike protein antibody cocktail.
Genomic analyses of the patient’s nasopharyngeal swabs confirmed an accelerated evolution of the virus through mutations in the spike gene and the receptor-binding domain.
In summary, these two case studies suggested an accelerated evolution and persistent shedding of the virus in conditions of immunodeficiency.
In particular, the first case highlighted the role of convalescent plasma in creating escape variants.
In fact, one study [292] exposed the SARS-CoV-2 virus to convalescent plasma in vitro repeatedly to see how much plasma was required to neutralize the virus.
The results of the first six exposures were similar, but they reported that after the seventh exposure (on day 45), the amount of plasma required began to increase.
In analyzing the viral variants present, they found that this viral escape was promoted by the sudden accumulation of mutations, especially in the receptor-binding domain (RBD) and N-terminal domain (NTD), that quickly rose in frequency.
By the thirteenth exposure (day 85), the virus had evolved three mutations and could not longer be neutralized by the plasma used.
Taken together, these observations suggest that evolutionary analyses of SARS-CoV-2 can provide crucial information about the conditions that promote resistance in SARS-CoV-2 and the kinetics of how resistance develops, information which will be important for understanding the implications of how vaccine regimens are designed and whether/when next-generation vaccines will be needed.
When variants occur, they can rise in frequency by chance or through an adaptive process that confers a competitive advantage to the virus.
Variants that had the D614G mutation in the spike glycoprotein seemed to spread faster.
However, it has been suggested that the mutation rose in frequency due to early chance events rather than by adaptive events [197].
Another mutation, Y453F, that occurred in the receptor binding domain of S, was first detected in mink; however, the transmission to humans has been established.
In mink, this mutation conferred an advantage by increasing the affinity towards ACE2 [293].
Similarly, N501Y mutation induces an increased affinity towards human ACE2 and has been involved in the dominance of B.1.1.7 by outcompeting other variants [275].
Therefore, genomic surveillance is essential to prevent the emergence of super-spreaders [294].
Emerging methods are being applied to this problem in an effort to understand which mutations are most likely to be of significant concern.
Novel machine learning methods were developed to predict the mutations in the sequence that promote viral escape.
While they preserve the pathogenicity of the virus, escape mutations change the virus’s sequence to evade detection by the immune system.
By using tools from natural language processing (NLP), viral escape was modeled as an NLP problem [295] where a modification makes a sentence grammatically correct but semantically different.
Therefore, language models of viruses could predict mutations that change the presentation of the virus to the immune system but preserve its infectivity.
3.6.3 Variants of concern and variants under surveillance
Viral replication naturally leads to the occurrence of mutations, and thus to genetic variation [296].
However, due to an intrinsic RNA proof-reading process in the SARS-CoV-2 virus, the pace of evolution of SARS-CoV-2 is moderate in comparison to other viruses [297].
The declaration of the first SARS-CoV-2 variant of concern (VOC) B.1.1.7 in December 2020 has attracted significant media attention.
While the B.1.1.7 lineage garnered attention in November 2020, two genomes of the lineage were detected as early as September 20th, 2020 from routine genomic data sampled in Kent (U.K.) by the COVID-19 Genomics UK Consortium (COG-UK).
The following day, a second B.1.1.7 genome was reported in greater London [197,298,299,300]
Since then, B.1.1.7 has spread across the UK and internationally, and it has now been detected in at least 62 countries [277], despite several countries imposing travel restrictions on travelers from the UK.
Of the twenty-three mutations that define B.1.1.7 from the original strain isolated in Wuhan (lineage A), fourteen are lineage-specific and three appear to be biologically consequential mutations associated with the spike protein, namely N501Y, P681H, and 69-70del [298,299].
The latter is a 6-bp deletion that leads to the loss of two amino acids and has consequences for immune recognition; it may, in conjunction with N501Y, be responsible for the increased transmissibility of the B.1.1.7 VOC due to changes in the RBD that increase binding affinity with ACE2 [281,298].
B.1.1.7 has increased transmissibility by up to 56%, leading to an R0 of approximately 1.4.
While there is also the possibility that this VOC may be associated with increased disease severity, there is currently insufficient evidence to draw any conclusions [274,275,300,301].
Other variants also express the 69-70del mutation [302,303], and public health officials in the United States and the UK have been able to use RT-PCR-based assays (ThermoFisher TaqPath COVID-19 assay) to identify sequences with this deletion because it occurs where the qPCR probe binds [300].
In the UK, B.1.1.7 is present in more than 97% of diagnostic tests that return negative for S-gene targets and positive for the other targets; thus, the frequency of S-gene target failure can be used as a proxy for the detection of B.1.1.7 [298; https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/950823/Variant_of_Concern_VOC_202012_01_Technical_Briefing_3_-England.pdf].
The FDA has highlighted that the performance of three diagnostic tests may be affected by the B.1.1.7 lineage because it could cause false negative tests [304].
While B.1.1.7 is currently the main VOC, other genetic variants not designated VOCs have been detected, including B.1.351 and P.1, both of which emerged independently [305].
B.1.351 was first detected in October 2020 in South Africa, was later detected in the EU on December 28th, 2020 and has now spread to at least 26 countries [282,306,307].
B.1.351 contains several mutations at the RBD including K417N, E484K, and N501Y.
While the biological significance of these mutations are still under investigation, it does appear that this lineage may be associated with increased transmissibility [308] due to the N501Y mutation [281,299].
The P.1 variant is a sublineage of the B.1.1.28 lineage that was first detected in Japan in samples obtained from four travelers from Brazil during a screening at a Tokyo airport on January 10, 2021 [309].
Shortly thereafter, it was established that there was a concentration of cases of the P.1 variant in Manaus, in the Amazonas State, Brazil.
In a small number of samples (n=31) sequenced in Manaus, 42% were identified as the P.1 variant as early as mid-December, but the variant seemed to be absent in genome surveillance testing prior to December [310].
To date, at least eight countries have detected the P.1 lineage [311].
While the majority of P.1 cases detected internationally have been linked to travel originating from Brazil, the UK has also reported evidence of community transmission detected via routine community sequencing [311,312].
P.1 has eight lineage-specific mutations along with three concerning spike protein mutations in the RBD, including K417T, E484K, and N501Y [308].
There have been multiple different SARS-CoV-2 lineages detected that have mostly been of no more clinical concern than the original devastating lineage originating in Wuhan [313].
However, the spotlight has been cast on other variants of unknown clinical relevance due to the increase of cases observed that have been associated with B.1.1.7 in particular.
Although early in its ascendency, B.1.427/429 is SARS-CoV-2 variants that was detected in California, USA and also known as CAL.20C [314].
It was first detected in July 2020 but was not detected again until October 2020.
In December 2020, B.1.427/429 accounted for ~24% of the total cases in Southern California and ~36% of total cases in the Los Angeles area.
B.1.427/429 has now been detected in New York and Washington, D.C., and some cases have been reported outside the USA in Oceania [314].
This variant is characterized by five key lineage-specific mutations (ORF1a: I4205V, ORF1b:D1183Y, S: S13I;W152C;L452R).
The latter spike mutation, L452R, is found in an area of the RBD known to resist monoclonal antibodies to the spike protein [315], and it is hypothesized that this mutation may resist polyclonal sera in convalescent patients or in individuals post-vaccination [314,316].
B.1.427/429 is not a designated VOC; however, further research is required to determine the implications of the mutations encoded in this genetic variant.
Another notable variant has recently been discovered in 35 patients in a Bavarian hospital in Germany; however, the sequencing data has not been published to date and it remains to be determined whether this variant is of any further concern [317].
There are several shared mutations and deletions between the three lineages, P.1, B.1.1.7, and B.1.315 and indeed other variants of SARS-CoV-2 that are under investigation [310].
For example, N501Y, which appears to have occurred independently in each of the three lineages.
E484K is present in both B.1.351 and P.1 [318].
The mutations N501Y and E484K are found in the RBD within the receptor-binding motif responsible for forming an interface with the ACE2 receptor, which seems to be consequential for ACE2 binding affinity [319].
Indeed, N501Y is associated with increased virulence and infectivity in mouse models [320].
E484K has also been associated with evasion from neutralizing antibodies [292,316,321].
The del69-70 (del:11288:9) is also shared between P.1 and B.1.1.7 and happens to be a common deletion found in the N terminal mutation of the spike protein.
This deletion has also been associated with several RBD mutations [281,299,322].
There is concern that mutations in the spike protein of variants may lead to clinical consequences for transmissibility, disease severity, re-infection, therapeutics, and vaccinations [292,316,323,324,325,326,327].
Vaccine producers are working to determine whether the vaccines are still effective against the novel genetic variants.
Moderna recently published data for their mRNA-1273 vaccine that showed no significant impact of neutralization against the B.1.1.7 variant upon vaccination in humans and non-human primates.
On the other hand, Moderna reported a reduced but significant neutralization against the B.1.351 variant upon vaccination [328].
Indeed, Pfizer–BioNTech reported that sera from twenty participants vaccinated with the BNT162b COVID-19 vaccine in previous clinical trials [329,330] elicited equivalent neutralizing titers against isogenic Y501 SARS-CoV-2 on an N501Y genetic background in vitro[331].
Another study has reported that the plasma neutralizing activity against SARS-CoV-2 variants encoding the combination of K417N:E484K:N501Y or E484K or N501Y was variably and significantly reduced in the sera of twenty participants who received either the Pfizer–BioNTech BNT162b (n = 6) vaccine or the Moderna’s mRNA-1273 vaccine (n =14) [332].
For now, the consensus appears to be that the FDA approved vaccines still seem to be effective against the genetic variants of SARS-CoV-2 and their accompanying mutations, albeit with a slightly lower neutralizing capacity [328,331,332,333].
Further research is required to discern the clinical, prophylactic, and therapeutic consequences of these genetic SARS-CoV-2 variants as the pandemic evolves.
3.7 Conclusions
As of October 2020 the SARS-CoV-2 virus remains a serious worldwide threat.
The scientific community has responded by rapidly collecting and disseminating information about the SARS-CoV-2 virus and the associated illness, COVID-19.
The rapid identification of the genomic sequence of the virus allowed for early contextualization of SARS-CoV-2 among other known respiratory viruses.
The pathogen is a coronavirus that is closely related to SARS-CoV-1, which caused the SARS pandemics of the early 2000s.
Knowing the phylogenetic context and genomic sequence of the virus then allowed for rapid insights into its structure and pathogenesis.
4 Diagnostics
4.1 Abstract
4.2 Importance
4.3 Introduction
Since the emergence of Severe acute respiratory syndrome-like coronavirus 2 (SARS-CoV-2) in late 2019, significant international concern has focused on how to manage the spread of the virus.
Identifying individuals who have contracted coronavirus disease 2019 (COVID-19) is crucial to slowing down the global pandemic.
Given the high transmissibility of SARS-CoV-2 and the potential for asymptomatic or presymptomatic individuals to be contagious [1], the development of rapid, reliable, and affordable methods to detect SARS-CoV-2 infection is vitally important.
For instance, test-trace-isolate procedures are a cornerstone of many nations’ efforts to control the outbreak [334,335,336].
The genetic sequence of the virus was first released by Chinese officials on January 10, 2020, and the first test to detect the virus was released about 13 days later [337].
This information is important to the development of diagnostic approaches using a variety of approaches.
There are two main classes of diagnostic tests: molecular tests, which can diagnose an active infection by identifying the presence of SARS-CoV-2, and serological tests, which can assess whether an individual was infected in the past via the presence or absence of antibodies against SARS-CoV-2.
Molecular tests are essential for identifying individuals for treatment and alerting their contacts to quarantine and be alert for possible symptoms.
Here, a patient sample is evaluated to determine the presence or absence of a viral target.
These techniques depend on knowledge of the viral genomic sequence for the development of targeted primers.
On the other hand, serological tests are useful for collecting population-level information for epidemiological analysis, as they can be used to estimate the extent of the infection in a given area.
Thus, they may be useful in efforts to better understand the percent of cases that manifest as severe versus mild and for guiding public health and economic decisions regarding resource allocation and counter-disease measures.
These tests typically detect the presence of antibodies in blood plasma samples.
In such enzyme-linked immunosorbent assay (ELISA) approaches, the detection of the antibodies depends on knowledge of a specific antibody-antigen interaction.
As the pandemic has evolved throughout 2020 and 2021, a variety of technological implementations have emerged within these two categories.
4.4 Molecular Tests
Molecular tests are used to identify distinct genomic subsequences of a viral molecule in a sample and thus to diagnose an active viral infection.
An important first step is identifying which biospecimens are likely to contain the virus in infected individuals and then acquiring these samples from the patient(s) to be tested.
Common sampling sources for molecular tests include nasopharyngeal cavity samples, such as throat washes, throat swabs, and saliva [338], and stool samples [339].
Once a sample from an appropriate source is acquired from a patient, molecular tests can utilize a number of different steps, described below, to analyze a sample and identify whether evidence of SARS-CoV-2 is present.
When testing for RNA viruses like SARS-CoV-2, pre-processing is necessary in order to create DNA from the RNA sample.
The DNA can then be amplified with PCR.
Some tests use the results of the PCR itself to determine whether the pathogen is present, but in other cases, it may be necessary to sequence the amplified DNA.
For sequencing, an additional pre-processing step, library preparation, therefore must be undertaken.
Library preparation is the process of preparing the sample for sequencing, typically by fragmenting the sequences and adding adapters [340].
In some cases, library preparation can involve other modifications of the sample, such as adding barcodes to identify a particular sample within the sequence data.
Barcoding can therefore be used to pool samples from multiple sources.
There are different reagents used for library preparation that are specific to identifying one or more target sections with PCR [341].
Sequential pattern matching is then used to identify unique subsequences of the virus, and if sufficient subsequences are found, the test is considered positive.
Therefore, tests that utilize sequencing require a number of additional molecular and analytical steps relative to tests that use PCR alone.
4.4.1 RT-PCR
Real-time polymerase chain reaction (RT-PCR) tests determine whether a target is present by measuring the rate of amplification during PCR compared to a standard.
When the target is RNA, such as in the case of RNA viruses, the RNA must be converted into complementary DNA during pre-processing.
The first test developed and validated for the detection of SARS-CoV-2 uses RT-PCR with reverse transcription [337] to detect several regions of the viral genome: the ORF1b of the RNA-dependent RNA polymerase (RdRP), the Envelope protein gene (E), and the Nucleocapsid protein gene (N).
The publication reporting this text was released on January 23, 2020, less than two weeks after the sequence of the virus was first reported [337].
In collaboration with several other labs in Europe and in China, the researchers confirmed the specificity of this test with respect to other coronaviruses against specimens from 297 patients infected with a broad range of respiratory agents.
Specifically this test utilizes two probes against RdRP, one of which is specific to SARS-CoV-2 [337].
Importantly, this assay was not found to return false positive results.
4.4.2 RT-qPCR
Another test was also announced during January 2020.
Chinese researchers developed a reverse transcriptase quantitative real-time PCR (RT-qPCR) test to identify two gene regions of the viral genome, ORF1b and N[342].
This assay was tested on samples from two COVID-19 patients and a panel of positive and negative controls consisting of RNA extracted from several cultured viruses.
The assay uses the N gene to screen patients, while the ORF1b gene region is used to confirm the infection [342].
In this case the test was designed to detect sequences conserved across sarbecoviruses, or viruses within the same subgenus as SARS-CoV-2.
Considering that SARS-CoV-1 and SARS-CoV-2 are the only sarbecoviruses currently known to infect humans, a positive test can be assumed to indicate that the patient is infected with SARS-CoV-2.
However, this test is not able to discriminate the genetics of viruses within the sarbecovirus clade.
4.4.2.1 dPCR
Digital PCR (dPCR) is a new generation of PCR technologies offering an alternative to traditional real-time quantitative PCR.
In dPCR, a sample is partitioned into thousands of compartments, such as nanodroplets (droplet dPCR or ddPCR) or nanowells, and a PCR reaction takes place in each compartment.
This design allows for a digital read-out where each partition is either positive or negative for the nucleic acid sequence being tested for, allowing for much higher throughput.
While dPCR equipment is not yet as common as that for RT-PCR, dPCR for DNA targets generally achieves higher sensitivity than other PCR technologies while maintaining high specificity, though sensitivity is slightly lower for RNA targets [343].
High sensitivity is particularly relevant for SARS-CoV-2 detection, since low viral load in clinical samples can lead to false negatives.
Suo et al. [344] performed a double-blind evaluation of ddPCR for SARS-CoV-2 detection on 57 samples, comprised by 43 samples from suspected positive patients and 14 from supposed convalescents, that had all tested negative for SARS-CoV-2 using RT-PCR.
Despite the initial negative results, 33 out of 35 (94.3%) patients were later clinically confirmed positive.
All of these individuals tested positive using ddPCR.
Additionally, of 14 supposed convalescents who had received two consecutive negative RT-PCR tests, nine (64.2%) tested positive for SARS-CoV-2 using ddPCR.
Two symptomatic patients tested negative with both RT-PCR and ddPCR, but were later clinically diagnosed positive, and 5 of the 14 suspected convalescents tested negative by ddPCR.
While this study did not provide a complete head-to-head comparison to RT-PCR in all aspects, e.g., no samples testing positive using RT-PCR were evaluated by ddPCR, the study shows the potential of dPCR for viral detection even in highly diluted samples.
A second study [345] confirmed that RT-ddPCR is able to detect SARS-CoV-2 at a lower threshold for viral load relative to RT-PCR.
This study analyzed 196 samples, including 103 samples from suspected patients, 77 from contacts and close contacts, and 16 from suspected convalescents, using both RT-qPCR and RT-ddPCR.
Of the 103 suspected patient samples, RT-qPCR identified 29 as positive, 25 as negative, and 49 as suspected.
Of the RT-qPCR negative or suspected samples, a total of 61 (19 negative and 42 suspected) were confirmed to be positive by RT-ddPCR.
All 103 of the suspected patients were later confirmed to be SARS-CoV-2 positive through a combination of symptom development and RT-qPCR resampling, indicating that RT-ddPCR improved the overall detection rate among these patients from 28.2% to 87.4%.
Of 77 patient samples from contacts and close contacts, 48 tested negative with both methods, and these patients were observed to remain healthy over a period of 14 days.
Within the remaining 29 patient samples, 12 tested positive, 1 negative, and 16 suspected with RT-qPCR.
Fifteen out of 16 suspected results and the negative result were overturned by positive RT-ddPCR results, decreasing the rate of suspected cases from 21% to 1%.
All 16 patients identified as positive by RT-ddPCR were subsequently determined, both clinically and through repeated sampling, to be positive for SARS-CoV-2.
Importantly, all samples that tested positive using RT-qPCR also tested positive using ddPCR.
Among the 16 convalescent patients, RT-qPCR identified 12 as positive, three as suspect, and one as negative, but RT-dPCR identified all 16 as positive.
This evidence further indicates that the lower limit of detection made possible by ddPCR may be useful for identifying when COVID-19 patients are cleared of the virus.
Overall, these studies suggest that ddPCR is a promising tool for overcoming the problem of false-negative SARS-CoV-2 testing.
4.4.3 Pooled and Automated PCR Testing
Due to limited supplies and the need for more tests, several labs have found ways to pool or otherwise strategically design tests to increase throughput.
The first such result came from Yelin et al. [346], who found they could pool up to 32 samples in a single qPCR run.
This was followed by larger-scale pooling with slightly different methods [347].
Although these approaches are also PCR based, they allow for more rapid scaling and higher efficiency for testing than the initial PCR-based methods developed.
Technology based on CRISPR (clustered regularly interspaced short palindromic repeats) has also been instrumental in scaling up testing protocols.
4.4.3.1 CRISPR-based Detection
Two CRISPR-associated nucleases, Cas12 and Cas13, have been used for nucleic acid detection.
Multiple assays exploiting these nucleases have emerged as potential diagnostic tools for the rapid detection of SARS-CoV-2 genetic material and therefore SARS-CoV-2 infection.
The SHERLOCK method (Specific High-sensitivity Enzymatic Reporter unLOCKing) from Sherlock Biosciences relies on Cas13a to discriminate between inputs that differ by a single nucleotide at very low concentrations [348].
The target RNA is amplified by RT-RPA and T7 transcription, and the amplified product activates Cas13a.
The nuclease then cleaves a reporter RNA, which liberates a fluorescent dye from a quencher.
Several groups have used the SHERLOCK method to detect SARS-CoV-2 viral RNA.
An early study reported that the method could detect 7.5 copies of viral RNA in all 10 replicates, 2.5 copies in 6 out of 10, and 1.25 copies in 2 out of 10 runs [349].
It also reported 100% specificity and sensitivity on 114 RNA samples from clinical respiratory samples (61 suspected cases, among which 52 were confirmed and nine were ruled out by metagenomic next-generation sequencing, 17 nCoV-/HCoV+ cases and 36 samples from healthy subjects), and a reaction turnaround time of 40 minutes.
A separate study screened four designs of SHERLOCK and extensively tested the best-performing assay.
They determined the limit of detection to be 10 copies/μl using both fluorescent and lateral flow detection [350].
Lateral flow test strips are simple to use and read, but there are limitations in terms of availability and cost per test.
Another group therefore proposed the CREST protocol (Cas13-based, Rugged, Equitable, Scalable Testing), which uses a P51 cardboard fluorescence visualizer powered by a 9-volt battery, for the detection of Cas13 activity instead of immunochromatography [351].
CREST can be run from RNA sample to result in approximately 2 hours, with no need for AC power or a dedicated facility and with minimal handling.
Testing was performed on 14 nasopharyngeal swabs.
CREST picked up the same positives as the CDC-recommended TaqMan assay with the exception of one borderline sample that displayed low-quality RNA.
This approach may therefore represent a rapid, accurate, and affordable procedure for detecting SARS-CoV-2.
The DETECTR method (DNA Endonuclease-Targeted CRISPR Trans Reporter) from Mammoth Biosciences involves purification of RNA extracted from patient specimens, amplification of extracted RNAs by loop-mediated amplification, which is a rapid, isothermal nucleic acid amplification technique, and application of their Cas12-based technology.
In this assay, guide RNAs (gRNAs) were designed to recognize portions of sequences corresponding to the SARS-CoV-2 genome, specifically the N2 and E regions [352].
In the presence of SARS-CoV-2 genetic material, sequence recognition by the gRNAs results in double-stranded DNA cleavage by Cas12, as well as cleavage of a single-stranded DNA molecular beacon.
The cleavage of this molecular beacon acts as a colorimetric reporter that is subsequently read out in a lateral flow assay and indicates the positive presence of SARS-CoV-2 genetic material and therefore SARS-CoV-2 infection.
The 40-minute assay is considered positive if there is detection of both the E and N genes or presumptive positive if there is detection of either of them.
The assay had 95% positive predictive agreement and 100% negative predictive agreement with the US Centers for Disease Control and Prevention SARS-CoV-2 real-time RT–PCR assay.
The estimated limit of detection was 10 copies per μl reaction, versus 1 copy per μl reaction for the CDC assay.
These results have been confirmed by other DETECTR approaches.
Using real-time recombinase polymerase amplification (RT-RPA) for amplification, another group detected 10 copies of synthetic SARS-CoV-2 RNA per μl of input within 60 minutes of RNA sample preparation in a proof-of-principle evaluation [353].
The DETECTR protocol was improved by combining RT-RPA and CRISPR-based detection in a one-pot reaction that incubates at a single temperature, and by using dual CRISPR RNAs (which increases sensitivity).
This new assay, known as All-In-One Dual CRISPR-Cas12a (AIOD-CRISPR), detected 4.6 copies of SARS-CoV-2 RNA per μl of input in 40 minutes [354].
Another single-tube, constant-temperature approach using Cas12b instead of Cas12a achieved a detection limit of 5 copies/μl in 40-60 minutes [355].
It was also reported that electric field gradients can be used to control and accelerate CRISPR assays by co-focusing Cas12-gRNA, reporters, and target [356].
The authors generated an appropriate electric field gradient using a selective ionic focusing technique known as isotachophoresis (ITP) implemented on a microfluidic chip.
They also used ITP for automated purification of target RNA from raw nasopharyngeal swab samples.
Combining this ITP purification with loop-mediated isothermal amplification, their ITP-enhanced assay to achieved detection of SARS-CoV-2 RNA (from raw sample to result) in 30 minutes.
There is an increasing body of evidence that CRISPR-based assays offer a practical solution for rapid, low-barrier testing in areas that are at greater risk of infection, such as airports and local community hospitals.
In the largest study to date, DETECTR was compared to RT-qPCR on 378 patient samples [357].
The authors reported a 95% reproducibility.
Both techniques were equally sensitive in detecting SARS-CoV-2.
Lateral flow strips showed a 100% correlation to the high-throughput DETECTR assay.
Importantly, DETECTR was 100% specific for SARS-CoV-2 and did not detect other human coronaviruses.
4.4.4 Limitations of Molecular Tests
Tests that identify SARS-CoV-2 using nucleic-acid-based technologies will identify only individuals with current infections and are not appropriate for identifying individuals who have recovered from a previous infection.
Within this category, different types of tests have different limitations.
For example, PCR-based test can be highly sensitive, but in high-throughput settings they can show several problems:
First of all, there is a risk of false-negative responses, which can present a significant problem to large-scale testing.
To reduce occurrence of false negatives, correct execution of the analysis is crucial [358].
Additionally, the emerging nature of the COVID-19 pandemic has introduced some challenges related to Uncertainty surrounding interactions between SARS-CoV-2 and its human hosts.
For example, viral shedding kinetics are still not well understood, but are expected to introduce a significant effect of timing of sample collection on test results [358].
Similarly, the type of specimen could also influence outcomes, as it is not clear which clinical samples are best for detecting the virus [358].
There are also significant practical and logistical concerns.
Much of the technology used for molecular tests is expensive, and while it might be available in major hospitals and/or diagnostic centers, it is often not available to smaller facilities [359].
At times during the first year of the pandemic, the availability of supplies for testing, including swabs and testing media, has also been limited [360].
Similarly, processing times can be long, and tests might take up to 4 days to return results [359].
Finally, with CRISPR-based testing strategies, the gRNA can recognize other interspersed sequences on the patient’s genome, false positives and a loss of specificity can occur.
As noted above, false negatives are a significant concern for several reason.
Importantly, clinical reports indicate that it is imperative to exercise caution when interpreting the results of molecular tests for SARS-CoV-2 because negative results do not necessarily mean a patient is virus-free [361].
4.5 Serological Tests
Although diagnostic tests based on the detection of genetic material can be quite sensitive, they provide information only about active infection, and therefore offer just a snapshot-in-time perspective on the spread of a disease.
Most importantly, they would not work on a patient who has fully recovered from the virus at the time of sample collection.
In this context, serological tests, which use serum to test for the presence of antibodies against SARS-CoV-2, are significantly more informative.
Serological tests can provide insight into population-level dynamics and can also offer a glimpse into the development of antibodies by individual patients during the course of a disease.
Therefore, they can be useful to developing strategies for the management of viral spread.
Furthermore, serological tests hold significant interest because of the possibility that they could provide information relevant to advancing economic recovery and allowing reopenings.
For instance, early in the course of the COVID-19 pandemic, it was hypothesized that people who had developed antibodies might be able to return to work [362], although this strategy would have relied on recovered individuals acquiring long-term immunity.
Some infectious agents can be controlled through “herd immunity”, which is when a critical mass within the population acquires immunity through vaccination and/or infection, preventing an infectious agent from spreading widely.
A simple SIR model predicts that to achieve the required level of exposure for herd immunity to be effective, at least (1-(1/R0)) fraction of the population must be immune or, equivalently, less than (1/R0) fraction of the population susceptible [243].
However, for SARS-CoV-2 and COVID-19, the R0 and mortality rates that have been observed suggest that relying on herd immunity without some combination of vaccines, proven treatment options, and strong non-pharmaceutical measures of prevention and control would likely result in a significant loss of life.
<~–Include some of this info somewhere?
Understanding the fundamental organization of the human immune response to viral threats is critical to understanding the varied response to SARS-CoV-2.
The human immune system utilizes a variety of innate and adaptive responses to protect against the pathogens it encounters.
The innate immune system consists of barriers, such as the skin, mucous secretions, neutrophils, macrophages, and dendritic cells.
It also includes cell-surface receptors that can recognize the molecular patterns of pathogens.
The adaptive immune system utilizes antigen-specific receptors that are expressed on B and T lymphocytes.
These components of the immune system typically act together; the innate response acts first, and the adaptive response begins to act several days after initial infection following the clonal expansion of T and B cells [35].
After a virus enters into a host cell, its antigen is presented by major histocompatibility complex 1 (MHC 1) molecules and is then recognized by cytotoxic T lymphocytes.
–>
4.5.1 Sustained Immunity to COVID-19
In the process of mounting a response to a pathogen, the immune system produces antibodies specific to the pathogen.
Understanding the acquisition and retention of antibodies is important both to the diagnosis of prior (inactive) infections and to the development of vaccines.
The two immunoglobulin classes most pertinent to these goals are immunoglobulin M (IgM), which are the first antibodies produced in response to an infection, and immunoglobulin G (IgG), which are the most abundant antibodies found in the blood.
Prior research is available about the development of antibodies to Severe acute respiratory syndrome-related coronavirus 1 (SARS-CoV-1) during the course of the associated disease, severe acute respiratory syndrome (SARS).
Following SARS infection, IgM and IgG antibodies were detected in the second week post-infection.
IgM titers peaked by the first month post-infection, and then declined to undetectable levels after day 180.
IgG titers peaked by day 60, and persisted in all donors through the two-year duration of study [363].
A two-year longitudinal study following convalesced SARS patients with a mean age of 29 found that IgG antibodies were detectable in all 56 patients surveyed for at least 16 months, and remained detectable in all but 4 (11.8%) of patients through the full two-year study period [364].
These results suggest that immunity to SARS-CoV-1 is sustained for at least a year.
The persistence of antibodies to SARS-CoV-2 remains under investigation.
Circulating antibody titers to other coronaviruses have been reported to decline significantly after 1 year [365].
Autopsies of lymph nodes and spleens from severe acute COVID-19 patients showed a loss of T follicular helper cells and germinal centers that may explain some of the impaired development of antibody responses [366].
An early study (initially released on medRxiv on February 25, 2020) presented a chemiluminescence immunoassay to a synthetic peptide derived from the amino acid sequence of the SARS-CoV-2 S protein [367].
This method was highly specific to SARS-CoV-2 and detected IgM in 57.2% and IgG in 71.4% and 57.2% of sera samples from 276 confirmed COVID-19 patients.
It reported that IgG could be detected within two days of the onset of fever but that IgM could not be detected any earlier, a pattern they compared to findings in another disease caused by a HCoV, Middle East respiratory syndrome (MERS).
Since then, several trials have reported the potential protective effect of antibodies in convalescent plasma obtained from recovered COVID-19 patients to treat critically ill COVID-19 patients [368,369,370].
Evidence to date suggests that sustained immunity to the SARS-CoV-2 virus remains for a period of at least 6 to 8 months [371,372,373,374].
Dan et al. assessed sustained immunity using 254 blood samples from 188 COVID-19 positive patients [372].
The samples were collected at various time points between 6 and 240 days post-symptom onset, meaning some patients were assessed longitudinally.
Of the samples, 43 were collected at least 6 months after symptom onset.
After 1 month, 98% of patients were seropositive for IgG to the spike protein, S.
Moreover, S IgG titers were stable and heterogeneous among patients over a period of 6 to 8 months post-symptom onset, with 90% of subjects seropositive at 6 months.
Similarly, at 6 to 8 months 88% of patients were seropositive for receptor binding domain (RBD) IgG and 90% were seropositive for SARS-CoV-2 neutralizing antibodies.
The findings of Dan et al. are in accordance with a study by Sherina et al. that examined 119 samples from 88 donors who had recovered from mild to severe cases of COVID-19 [374].
They observed a relatively stable level of IgG and plasma neutralizing antibodies up to 6 months post diagnosis.
Significantly lower but considerable levels of anti-SARS-CoV-2 IgG antibodies were still present in 80% of samples obtained 6-8 months post symptom-onset.
A study by Gaebler et al. also found that titers of IgM and IgG antibodies against the RBD decreased from 1.3 to 6.2 months post infection in a study of 87 individuals [375].
However, the decline of IgA (15%) activity was less pronounced than that of IgM (53%) or IgG (32%).
It was noted that higher levels of anti-RBD IgG and anti-N total antibodies were detected in individuals that reported persistent post-acute symptoms at both study visits.
Moreover, plasma neutralizing activity decreased five-fold between 1.3 and 6.2 months in an assay of HIV-1 virus pseudotyped with SARS-CoV-2 S protein, and this neutralizing activity was directly correlated with IgG anti-RBD titers [375].
These findings are in accordance with other studies that show that the majority of seroconverters have detectable, albeit decreasing, levels of neutralizing antibodies at least 3-6 months post infection [376,377,378].
Determining the potency of anti-RBD antibodies early in the course of an infection may be important moving forward, as their neutralizing potency may be prognostic for disease severity and survival [379].
The development of memory B cells and memory T cells has also been assessed in several studies.
Dan et al. showed that SARS-CoV-2 S-specific memory B cell levels rose steadily over the first 120 days following symptom onset [372].
RBD-specific memory B cells had been detected in COVID-19 patients 90 days post-symptom onset in previous studies [373,380], and this study confirms these finding and shows that levels of these cells increased over the 4-5 months post-symptom onset [372].
Gaebler et al. have shown that memory B cells specific to the RBD remain unaltered and exhibit clonal turnover and antibody sequence evolution 6 months post infection, indicative of prolonged germinal cell reactions [375].
The same study showed that antibodies expressed by these memory B cells have resistance to RBD mutations, greater somatic hypermutations, and increased potency, which the authors suggest might be evidence of continued evolution of humoral immunity [375].
Indeed, Wheatley et al. showed that S-specific IgG+ memory B cells consistently increase over time and by 4 months comprise approximately 0.8% of all IgG+ memory B cells, which may indicate cellular immune memory to even mild-to-moderate COVID-19 infection [378].
Dan et al. showed that N-specific memory B cells steadily increased up to 4-5 months post-symptom onset [372].
SARS-CoV-2 memory CD8+ T cells were also detected in 70% of 169 COVID-19 patients after 1 month [372], which is consistent with previous research [381].
However, SARS-CoV-2 memory CD8+ T cells were slightly decreased (50%) 6 months post-symptom onset.
In this same subset of COVID-19 patients, 93% of subjects had detectable levels of SARS-CoV-2 memory CD4+ T cells, of which 42% had more than 1% SARS-CoV-2-specific CD4+ T cells.
At 6 months, 92% of patients were positive for SARS-CoV-2 memory CD4+ T cells.
Indeed, the abundance of S-specific memory CD4+ T cells over time was similar to that of SARS-CoV-2-specific CD4+ T cells overall [372].
T cell immunity to SARS-CoV-2 at 6 to 8 months following symptom onset has also been confirmed by other studies [374,382,383].
In another study, T cell reactivity to SARS-CoV-2 epitopes was also detected in some individuals never been exposed to SARS-CoV-2.
This finding suggests the potential for cross-reactive T cell recognition between SARS-CoV-2 and pre-existing circulating HCoV that are responsible for the “common cold” [381], but further research is required.
Concerns have also been raised that immunity may wane over time.
Several reported cases of reinfection have been confirmed via genomic analysis that revealed distinct variants of SARS-CoV-2 within a single patient [384,385,386].
Further research is required to determine the full extent to which sustained immunity can be and is typically achieved following SARS-CoV-2 infection.
Furthermore, it is unclear whether previous infection may carry repercussions in terms of disease severity for patients [387] and what implications the possibility of reinfection holds for vaccine development and the long-term efficacy of vaccines [388,389].
4.5.2 Current Approaches
Several countries are now focused on implementing antibody tests, and in the United States, the FDA recently approved a serological test by Cellex for use under emergency conditions [390].
Specifically, the Cellex qSARS-CoV-2 IgG/IgM Rapid Test is a chromatographic immunoassay designed to qualitatively detect IgM and IgG antibodies against SARS-CoV-2 in the plasma (from a blood sample) of patients suspected to have developed the SARS-CoV-2 infection [390].
Such tests illuminate the progression of viral disease, as IgM are the first antibodies produced by the body and indicate that the infection is active.
Once the body has responded to the infection, IgG are produced and gradually replace IgM, indicating that the body has developed immunogenic memory [391].
The Cellex test cassette contains a pad of SARS-CoV-2 antigens and a nitrocellulose strip with lines for each of IgG and IgM, as well as a control (goat IgG) [390].
In a specimen that contains antibodies against the SARS-CoV-2 antigen, the antibodies will bind to the strip and be captured by the IgM and/or IgG line(s), resulting in a change of color [390].
With this particular assay, results can be read within 15 to 20 minutes [390].
Other research groups, such as the Krammer lab of the Icahn School of Medicine at Mount Sinai, proposed an ELISA test that detects IgG and IgM that react against the RBD of the spike proteins (S) of the virus [392].
The authors are now working to get the assay into clinical use [393].
4.5.3 Limitations of Serological Tests
Like molecular tests, serological tests carry a number of limitations that influence their utility in different situations.
Importantly, false positives can occur due to cross-reactivity with other antibodies according to the clinical condition of the patient [390].
Therefore, such tests must be used in combination with RNA detection tests if intended for diagnostic purposes, and while serological tests may be of interest to individuals who wish to confirm they were infected with SARS-CoV-2 in the past, their potential for false positives means that they are not currently recommended for this use.
Due to the long incubation times and delayed immune responses of infected patients, serological tests are insufficiently sensitive for a diagnosis in the early stages of an infection.
The limitations due to timing make serological tests far less useful for enabling test-and-trace strategies.
4.6 Possible Alternatives to Current Practices for Identifying Active Cases
COVID-19 can present with symptoms similar to other types of pneumonia, and symptoms can vary widely among COVID-19 patients; therefore, clinical presentation is often insufficient as a sole diagnostic criterion.
In addition, identifying and isolating mild or asymptomatic cases is critical to efforts to manage outbreaks.
Even among mildly symptomatic patients, a predictive model based on clinical symptoms had a sensitivity of only 56% and a specificity of 91% [394].
More problematic is that clinical symptom-based tests are only able to identify already symptomatic cases, not presymptomatic or asymptomatic cases.
They may still be important for clinical practice, and for reducing tests needed for patients deemed unlikely to have COVID-19.
Similarly, X-ray diagnostics have been reported to have high sensitivity but low specificity in some studies [395].
Other studies have shown that specificity varies between radiologists [396], though the sensitivity reported here was lower than that published in the previous paper.
However, preliminary machine-learning results have shown far higher sensitivity and specificity from analyzing chest X-rays than was possible with clinical examination [397].
X-ray tests with machine learning can potentially detect asymptomatic or presymptomatic infections that show lung manifestations.
This approach would still not recognize entirely asymptomatic cases.
Given the above, the widespread use of X-ray tests on otherwise healthy adults is likely inadvisable.
4.7 Strategies and Considerations for Determining Whom to Test
Early in the COVID-19 pandemic, testing was typically limited to individuals considered high risk for developing serious illness [398].
This approach often involved limiting testing to people with severe symptoms and people showing mild symptoms that had been in contact with a person who had tested positive.
Individuals who were asymptomatic (i.e., potential spreaders) and individuals who were able to recover at home were therefore often unaware of their status.
However, this method of testing administration misses a high proportion of infections and does not allow for test-and-trace methods to be used.
For instance, a recent study from Imperial College estimates that in Italy, the true number of infections was around 5.9 million in a total population ~60 million, compared to the 70,000 detected as of March 28th [259].
Another analysis, which examined New York state, indicated that as of May 2020, approximately 300,000 cases had been reported in a total population of approximately 20 million [399].
This corresponded to ~1.5% of the population, but ~12% of individuals sampled statewide were estimated as positive through antibody tests (along with indications of spatial heterogeneity at higher resolution) [399].
Technological advancements that facilitate widespread, rapid testing will therefore improve the potential to accurately assess the rate of infection and aid in controlling the virus’ spread.
4.8 Conclusions
Major advancements have been made in identifying diagnostic approaches.
The development of diagnostic technologies have been rapid, beginning with the release of the SARS-CoV-2 viral genome sequence in January.
As of October 2020, a range of diagnostic tests have become available.
One class of tests uses PCR (RT-PCR or RT-qPCR) to assess the presence of SARS-CoV-2 RNA, while another typically uses ELISA to test for the presence of antibodies to SARS-CoV-2.
The former approach is useful for identifying active infections, while the latter measures hallmarks of the immune response and therefore can detect either active infections or immunity gained from prior infection.
Combining these tests leads to extremely accurate detection of SARS-CoV-2 infection (98.6%), but when used alone, PCR-based tests are recommended before 5.5 days after the onset of the illness and antibody tests after 5.5 days [400].
Other strategies for testing can also influence the tests’ accuracy, such as the use of nasopharyngeal swabs versus bronchoalveolar lavage fluid [400], which allow for trade-offs between patient’s comfort and test sensitivity.
Additionally, technologies such as digital PCR may allow for scale-up in the throughput of diagnostic testing, facilitating widespread testing.
One major question that remains is whether people who recover from SARS-CoV-2 develop sustained immunity, and over what period this immunity is expected to last.
Some reports have suggested that some patients may develop COVID-19 reinfections (e.g., [384]), but the rates of reinfection are currently unknown.
Serologic testing combined with PCR testing will be critical to confirming purported cases of reinfection and to identifying the duration over which immunity is retained and to understanding reinfection risks.
5 Identification and Development of Therapeutics for COVID-19
5.1 Abstract
After emerging in China in late 2019, the novel Severe acute respiratory syndrome-like coronavirus 2 (SARS-CoV-2) spread worldwide and as of early 2021, continues to significantly impact most countries.
Only a small number of coronaviruses are known to infect humans, and only two are associated with the severe outcomes associated with SARS-CoV-2: Severe acute respiratory syndrome-related coronavirus, a closely related species of SARS-CoV-2 that emerged in 2002, and Middle East respiratory syndrome-related coronavirus, which emerged in 2012.
Both of these previous epidemics were controlled fairly rapidly through public health measures, and no vaccines or robust therapeutic interventions were identified.
However, previous insights into the immune response to coronaviruses gained during the outbreaks of severe acute respiratory syndrome (SARS) and Middle East respiratory syndrome (MERS) have proved beneficial to identifying approaches to the treatment and prophylaxis of novel coronavirus disease 2019 (COVID-19).
A number of potential therapeutics against SARS-CoV-2 and the resultant COVID-19 illness were rapidly identified, leading to a large number of clinical trials investigating a variety of possible therapeutic approaches being initiated early on in the pandemic.
As a result, a small number of therapeutics have already been authorized by regulatory agencies such as the Food and Drug Administration (FDA) in the United States, and many other therapeutics remain under investigation.
Here, we describe a range of approaches for the treatment of COVID-19, along with their proposed mechanisms of action and the current status of clinical investigation into each candidate.
The status of these investigations will continue to evolve, and this review will be updated as progress is made.
5.2 Importance
The COVID-19 pandemic is a rapidly evolving crisis.
With the worldwide scientific community shifting focus onto the SARS-CoV-2 virus and the disease it causes, a large number of possible pharmaceutical approaches for treatment and prevention have been proposed.
What is known about each of these potential interventions evolved rapidly throughout 2020 and early 2021.
In March 2020, we began monitoring a range of candidates and have continued to update this manuscript as new information has become available.
Some therapeutics have been supported, others have been revealed to be unlikely to confer any therapeutic benefits, and most require more data before a conclusion can be drawn.
This rapidly changing area of research provides important insight into how the ongoing pandemic can be managed and also demonstrates the power of interdisciplinary collaboration to rapidly understand a virus and match its characteristics with existing or novel pharmaceuticals.
5.3 Introduction
The novel coronavirus Severe acute respiratory syndrome-related coronavirus 2 (SARS-CoV-2) emerged in late 2019 and quickly precipitated the worldwide spread of novel coronavirus disease 2019 (COVID-19).
COVID-19 is associated with symptoms ranging from none (asymptomatic) to mild to severe, with approximately 2% of patients dying from COVID-19-related complications, such as acute respiratory disease syndrome (ARDS) [1].
The virus is likely spread between people primarily by droplets, with the role of contact and aerosol transmission still in question [214,215].
As a result, public health guidelines have been critical to efforts to control the spread of the virus.
However, as of early 2021, COVID-19 remains a significant worldwide concern (Figure 2), with cases in some places surging far above the numbers reported during the initial outbreak in early 2020.
Due to the continued threat of the virus and the severity of the disease, the identification and development of prophylactic and therapeutic interventions have emerged as significant international priorities.
Both approaches hold valuable potential for controlling the impact of the disease.
Prophylactics bolster immunity to prevent an individual from contracting a disease, whereas therapeutics treat a disease in individuals who have already been infected.
While a number of vaccines have recently been developed, approved, or are currently being evaluated by the US Food and Drug Administration and other regional and international bodies, vaccination programs only started being rolled out at the very end of 2020 and beginning of 2021, meaning that treatments that palliate symptoms and prevent the most severe outcomes have been necessary for much of 2020 and will continue to be necessary for the foreseeable future [401].
Fortunately, prior developments during other recent pandemics, especially those caused by human coronaviruses (HCoV), have provided a number of hypotheses guiding a biomedical approach to the novel coronavirus infection.
2,783,800 COVID-19 deaths had been reported worldwide as of March 28, 2021 (Figure 2).
Figure 2:Cumulative global COVID-19 deaths since January 22, 2020.
Data are from the COVID-19 Data Repository by the Center for Systems Science and Engineering at Johns Hopkins University [402].
5.3.1 Lessons from Prior HCoV Outbreaks
SARS-CoV-2’s rapid shift from an unknown virus to a significant worldwide threat closely parallels the emergence of Severe acute respiratory syndrome-related coronavirus (SARS-CoV-1).
The first documented case of COVID-19 was reported in Wuhan, China in November 2019, and the disease quickly spread worldwide during the early months of 2020.
Similarly, the first case of SARS was reported in November 2002 in the Guangdong Province of China, and it spread within China and then into several countries across continents over the following months [31,33].
In fact, genome sequencing quickly revealed the virus causing COVID-19 to be a novel betacoronavirus closely related to SARS-CoV-1 [13].
There are many similarities but also some differences in the characteristics of the two viruses that determine how they spread.
SARS-CoV-1 infection is severe, with an estimated death rate for SARS of 9.5% [33], while estimates of the death rate associated with COVID-19 are much lower, at approximately 2% [1].
SARS-CoV-1 is highly contagious via droplet transmission and has a basic reproduction number (R0) of 4 (i.e., each person infected was estimated to infect four other people) [33].
SARS-CoV-2 also appears to be spread primarily by droplet transmission [214,215], and most estimates of its R0 fall between 2.5 and 3 [1].
However, the 17-year difference in the timing of these two outbreaks has led to some major differences in the tools available for the international community’s response.
At the time that SARS-CoV-1 emerged, no new HCoV had been identified in almost 40 years [31].
The identity of the virus underlying the SARS disease remained unknown until April of 2003, when the SARS-CoV-1 virus was characterized through a worldwide scientific effort spearheaded by the World Health Organization (WHO) [31].
In contrast, the SARS-CoV-2 genomic sequence was released on January 3, 2020 [13], only days after the international community became aware of the novel pneumonia-like illness now known as COVID-19.
While SARS-CoV-1 belonged to a distinct lineage from the two other HCoVs known at the time of its discovery [33], SARS-CoV-2 is closely related to SARS-CoV-1 and a more distant relative of another HCoV characterized in 2012, Middle East respiratory syndrome-related coronavirus[16,403].
Despite their phylogenetic similarity, SARS-CoV-2 emerged under very different circumstances than SARS-CoV-1 in terms of scientific knowledge about HCoVs.
The trajectories of the pandemics associated with each of the viruses have also diverged significantly.
By July 2003, the SARS outbreak was officially determined to be under control, with the success credited to infection management practices such as mask wearing [31].
In contrast, Middle East respiratory syndrome (MERS) is still circulating and remains a concern; although the fatality rate is very high at almost 35%, the disease is much less easily transmitted, as its R0 has been estimated to be 1 [33].
The low R0 in combination with public health practices allowed for its spread to be contained [33].
Neither of these trajectories are comparable to that of SARS-CoV-2, which remains a serious threat worldwide more than a year after the first cases of COVID-19 emerged.
Current results suggest that pharmaceutical interventions for COVID-19 appear to be more successful than the previous efforts to develop therapeutics for SARS and MERS.
Care for SARS and MERS patients prioritized supportive care and symptom management [33].
To the extent that clinical treatments for SARS and MERS were explored, there is generally a lack of evidence supporting their efficacy.
For example, Ribavirin is an antiviral that was often used in combination with corticosteroids and sometimes interferon (IFN) medications to treat SARS and MERS [31], but its effects have been found to be inconclusive in retrospective and in vitro analyses of SARS and the SARS-CoV-1 virus, respectively [31].
IFNs and Ribavirin have shown promise in in vitro analyses of MERS, but their clinical effectiveness remains unknown [31].
Therefore, only limited pharmaceutical advances from prior HCoV outbreaks can be adopted to COVID-19.
Importantly, though, prior analyses of the virological and pathogenic properties of SARS-CoV-1 and Middle East respiratory syndrome-related coronavirus (MERS-CoV) have provided a strong foundation for the development of hypotheses about SARS-CoV-2 that have served to accelerated the development and identification of potential therapeutic approaches.
Even during the early months of the pandemic, a number of studies have emerged investigating the potential effects of drugs with mechanisms suggested based on prior understanding of coronavirus-host interactions.
Initially, these were primarily observational studies, which simply compare groups of patients who did and did not receive a treatment to determine whether it may have an effect.
However, these studies are subject to confounding, and randomized controlled trials are the standard means of assessing the effects of medications.
In randomized controlled trials, patients are prospectively and randomly assigned to treatment conditions, allowing for much stronger interpretations to be drawn; however, data from these trials takes much longer to collect.
Both have proven to be important sources of information during the COVID-19 crisis, but as more results are released from randomized controlled trials, more definitive answers are becoming available about various proposed therapeutics.
5.3.2 Overview of Therapeutic Approaches
Therapeutic approaches to the current pandemic can utilize two potential avenues: they can reduce the symptoms that are harmful to COVID-19 patients, or they can directly target the virus to hinder the spread of infection.
The goal of the former is to reduce the severity and risks of an active infection, while for the latter, it is to inhibit the replication of the virus once an individual is infected.
A variety of symptom profiles with a range of severity are associated with COVID-19, many of which are not life-threatening.
A study of COVID-19 patients in a hospital in Berlin, Germany found that the symptoms associated with the highest risk of death included infection-related symptoms, such as sepsis, respiratory symptoms such as ARDS, and cardiovascular failure or pulmonary embolism [404].
Therapeutics that reduce the risks associated with these severe outcomes hold particular potential to reduce the pandemic death toll, while therapeutics that directly target the virus itself would hold the potential to prevent people infected with SARS-CoV-2 from developing potentially damaging symptoms.
The treatments in this latter category typically fall into the broad category of antivirals.
Antiviral therapies hinder the spread of a virus within the host, rather than destroying existing copies of the virus, and these drugs can vary in their specificity to a narrow or broad range of viral targets.
For both categories, uncertainty often surrounds the treatments’ exact mechanisms of action, as most therapies have secondary or off-target effects.
A large number of clinical trials investigating a range of possible therapeutics for COVID-19 are currently in progress or have already been completed (Figure 3).
The purpose of this review is to critically appraise the literature surrounding a subset of clinical trials and to evaluate a range of approaches to repurpose existing or develop novel approaches to the mitigation and treatment of coronavirus infections.
The treatments evaluated here are classified according to their biological properties, specifically whether they are biologics (produced from components of organisms) or small molecules.
Small molecule drugs include drugs targeted at viral particles, drugs targeted at host proteins, and broad spectrum pharmaceuticals, while biologics include antibodies and interferons.
As we cover vaccines in a separate manuscript [401], we only consider therapies for the prevention of SARS-CoV-2 infection or COVID-19 in a very limited context here, for example when a drug is studied to see whether it can prevent infection or disease in recently-exposed individuals.
As results become available from additional clinical trials, we will continue to update this manuscript to keep pace with the current understanding of which therapeutics may be effective against SARS-CoV-2 or COVID-19.
Figure 3:COVID-19 clinical trials.
There are 6,417 COVID-19 clinical trials of which 3,706 are interventional.
The study types include only types used in at least five trials.
Interventional trials only are analyzed in the figures depicting status, phase, and intervention.
Of the interventional trials, 98 trials had reported results as of November 9, 2020.
Recruitment status and trial phase are shown only for interventional trials in which the status or phase is recorded.
The common interventions are all interventions used in at least ten trials.
Combinations of interventions, such as Hydroxychloroquine + Azithromycin, are tallied separately from the individual interventions.
Trials data are from the University of Oxford Evidence-Based Medicine Data Lab’s COVID-19 TrialsTracker [405].
5.4 Small Molecule Drugs
Small molecules are synthesized compounds of low molecular weight, typically less than 1 kilodalton (kDa) [406].
Small-molecule pharmaceutical agents have been a backbone of drug development since the discovery of penicillin in the early twentieth century [407].
It and other antibiotics have long been among the best known applications of small molecules to therapeutics, but biotechnological developments such as the prediction of protein-protein interactions have facilitated advances in precise targeting of specific structures using small molecules [407].
Small molecule drugs today encompass a wide range of therapeutics beyond antibiotics, including antivirals, protein inhibitors, and many broad-spectrum pharmaceuticals.
5.4.1 Small Molecule Antivirals
Antiviral drugs against SARS-CoV-2 are designed to inhibit replication of a virus within an epithelial host cell.
This process requires inhibiting the replication cycle of a virus by disrupting one of six fundamental steps [408] (Figure 4).
In the first of these steps, the virus attaches to and enters the host cell through endocytosis.
Then the virus undergoes uncoating, which is classically defined as the release of viral contents into the host cell.
Next, the viral genetic material enters the nucleus where it gets replicated during the biosynthesis stage.
During the assembly stage, viral proteins are translated, allowing new viral particles to be assembled.
In the final step new viruses are released into the extracellular environment.
Many antiviral drugs are designed to inhibit the replication of viral genetic material during the biosynthesis step.
Unlike DNA viruses, which can use the host enzymes to propagate themselves, RNA viruses like SARS-CoV-2 depend on their own polymerase, the RNA-dependent RNA polymerase (RdRP), for replication [409,410].
Targeting the RdRP is therefore an effective strategy for antivirals against RNA viruses and is the proposed mechanism underlying the treatment of SARS and MERS with Ribavirin [411].
However, although antivirals are designed to target a virus, they can also impact other processes in the host and may have unintended effects.
Therefore, these therapeutics must be evaluated for both efficacy and safety.
5.4.1.1 Nucleoside and Nucleotide Analogs
5.4.1.1.1 Favipiravir
Favipiravir (Avigan), also known as T-705, was discovered by Toyama Chemical Co., Ltd. [412].
The drug was found to be effective at blocking viral amplification in several influenza subtypes as well as other RNA viruses, such as Flaviviridae and Picornaviridae, through a reduction in plaque formation [413] and viral replication in Madin-Darby canine kidney cells [414].
Furthermore, inoculation of mice with favipiravir was shown to increase survival of influenza infections [413,414].
In 2014, the drug was approved in Japan for the treatment of influenza that was resistant to conventional treatments like neuraminidase inhibitors [415].
Favipiravir (6-fluoro-3-hydroxy-2-pyrazinecarboxamide) acts as a purine and purine nucleoside analogue that inhibits viral RNA polymerase in a dose-dependent manner across a range of RNA viruses, including influenza viruses [416,417,418,419,420].
Nucleotides and nucleosides are the natural building blocks for RNA synthesis.
Because of this, modifications to nucleotides and nucleosides can disrupt key processes including replication [421].
Biochemical experiments showed that favipiravir was recognized as a purine nucleoside analogue and incorporated into the viral RNA template.
A single incorporation does not influence RNA transcription; however, multiple events of incorporation lead to the arrest of RNA synthesis [422].
Evidence for T-705 inhibiting viral RNA polymerase are based on time-of-drug addition studies that found that viral loads were reduced with the addition of favipiravir in early times post-infection [416,419,420].
The effectiveness of favipiravir for treating patients with COVID-19 is currently under investigation.
An open-label, nonrandomized, before-after controlled study was recently conducted [423].
The study included 80 COVID-19 patients (35 treated with favipiravir, 45 control) from the isolation ward of the National Clinical Research Center for Infectious Diseases (The Third People’s Hospital of Shenzhen), Shenzhen, China.
The patients in the control group were treated with other antivirals, such as lopinavir and ritonavir.
It should be noted that although the control patients received antivirals, two subsequent large-scale analyses, the WHO Solidarity trial and the Randomized Evaluation of COVID-19 Therapy (RECOVERY) trial, identified no effect of lopinavir or of a lopinavir-ritonavir combination, respectively, on the metrics of COVID-19-related mortality that each assessed [424,425,426].
Treatment was applied on days 2-14; treatment stopped either when viral clearance was confirmed or at day 14.
The efficacy of the treatment was measured by, first, the time until viral clearance using Kaplan-Meier survival curves, and, second, the improvement rate of chest computed tomography (CT) scans on day 14 after treatment.
The study found that favipiravir increased the speed of recovery, measured as viral clearance from the patient by RT-PCR, with patients receiving favipiravir recovering in four days compared to 11 days for patients receiving antivirals such as lopinavir and ritonavir.
Additionally, the lung CT scans of patients treated with favipiravir showed significantly higher improvement rates (91%) on day 14 compared to control patients (62%, p = 0.004).
However, there were adverse side effects in 4 (11%) favipiravir-treated patients and 25 (56%) control patients.
The adverse side effects included diarrhea, vomiting, nausea, rash, and liver and kidney injury.
Despite the study reporting clinical improvement in favipiravir-treated patients, several study design issues are problematic and lower confidence in the overall conclusions.
For example, the study was neither randomized nor blinded.
Moreover, the selection of patients did not take into consideration important factors such as previous clinical conditions or sex, and there was no age categorization.
Additionally, it should be noted that this study was temporarily retracted and then restored without an explanation [427].
In late 2020 and early 2021, the first randomized controlled trials of favipiravir for the treatment of COVID-19 released results [428,429,430].
The first [428] used a randomized, controlled, open-label design to compare two drugs, favipiravir and baloxavir marboxil, to standard of care (SOC) alone.
Here, SOC included antivirals such as lopinavir/ritonavir and was administered to all patients.
The primary endpoint analyzed was viral clearance at day 14.
The sample size for this study was very small, with 29 total patients enrolled, and no significant effect of the treatments was found for the primary or any of the secondary outcomes analyzed, which included mortality.
The second study [429] was larger, with 96 patients enrolled, and included only individuals with mild to moderate symptoms who were randomized into two groups: one receiving chloroquine (CQ) in addition to SOC, and the other receiving favipiravir in addition to SOC.
This study reported a non-significant trend for patients receiving favipiravir to have a shorter hospital stay (13.29 days compared to 15.89 for CQ, p = 0.06) and less likelihood of progressing to mechanical ventilation (p = 0.118) or to an oxygen saturation < 90% (p = 0.129).
These results, combined with the fact that favipiravir was being compared to CQ, which is now widely understood to be ineffective for treating COVID-19, thus do not suggest that favipiravir was likely to have had a strong effect on these outcomes.
On the other hand, another trial of 60 patients reported a significant effect of favipiravir on viral clearance at four days (a secondary endpoint), but not at 10 days (the primary endpoint) [430].
This study, as well as a prior study of favipiravir [431], also reported that the drug was generally well-tolerated.
Thus, in combination, these small studies suggest that the effects of favipiravir as a treatment for COVID-19 cannot be determined based on the available evidence, but additionally, none raise major concerns about the safety profile of the drug.
5.4.1.1.2 Remdesivir
Remdesivir (GS-5734) is an intravenous antiviral that was proposed by Gilead Sciences as a possible treatment for Ebola virus disease (EVD).
At the outset of the COVID-19 pandemic, it did not have any have any FDA-approved use.
However, on May 1, 2020, the FDA issued an Emergency Use Authorization (EUA) for remdesivir for the treatment of hospitalized COVID-19 patients [432].
The EUA was based on information from two clinical trials, NCT04280705 and NCT04292899 [433,434,435,436].
Remdesivir is metabolized to GS-441524, an adenosine analog that inhibits a broad range of polymerases and then evades exonuclease repair, causing chain termination [437,438,439].
A clinical trial in the Democratic Republic of Congo found some evidence of effectiveness against EVD, but two antibody preparations were found to be more effective, and remdesivir was not pursued [440].
Remdesivir also inhibits polymerase and replication of the coronaviruses MERS-CoV and SARS-CoV-1 in cell culture assays with submicromolar IC50s [441].
It has also been found to inhibit SARS-CoV-2, showing synergy with CQ in vitro[439].
Remdesivir was first used on some COVID-19 patients under compassionate use guidelines [442,443].
All were in late stages of COVID-19 infection, and initial reports were inconclusive about the drug’s efficacy.
Gilead Sciences, the maker of remdesivir, led a recent publication that reported outcomes for compassionate use of the drug in 61 patients hospitalized with confirmed COVID-19.
Here, 200 mg of remdesivir was administered intravenously on day 1, followed by a further 100 mg/day for 9 days [436].
There were significant issues with the study design, or lack thereof.
There was no randomized control group.
The inclusion criteria were variable: some patients only required low doses of oxygen, while others required ventilation.
The study included many sites, potentially with variable inclusion criteria and treatment protocols.
The patients analyzed had mixed demographics.
There was a short follow-up period of investigation.
Eight patients were excluded from the analysis mainly due to missing post-baseline information; thus, their health was unaccounted for.
Therefore, even though the study reported clinical improvement in 68% of the 53 patients ultimately evaluated, due to the significant issues with study design, it could not be determined whether treatment with remdesivir had an effect or whether these patients would have recovered regardless of treatment.
Another study comparing 5- and 10-day treatment regimens reported similar results but was also limited because of the lack of a placebo control [445].
These studies did not alter the understanding of the efficacy of remdesivir in treating COVID-19, but the encouraging results provided motivation for placebo-controlled studies.
Remdesivir was later tested in a double-blind placebo-controlled phase 3 clinical trial performed at 60 trial sites, 45 of which were in the United States [434,435].
The trial recruited 1,062 patients and randomly assigned them to placebo treatment or treatment with remdesivir.
Patients were stratified for randomization based on site and the severity of disease presentation at baseline [435].
The treatment was 200 mg on day 1, followed by 100 mg on days 2 through 10.
Data was analyzed from a total of 1,059 patients who completed the 29-day course of the trial, with 517 assigned to remdesivir and 508 to placebo [435].
The two groups were well matched demographically and clinically at baseline.
Those who received remdesivir had a median recovery time of 10 days, as compared with 15 days in those who received placebo (rate ratio for recovery, 1.29; 95% confidence interval (CI), 1.12 to 1.49; p < 0.001).
The Kaplan-Meier estimates of mortality by 14 days were 6.7% with remdesivir and 11.9% with placebo, with a hazard ratio (HR) for death of 0.55 and a 95% CI of 0.36 to 0.83, and at day 29, remdesivir corresponded to 11.4% and the placebo to 15.2% (HR: 0.73; 95% CI, 0.52 to 1.03).
Serious adverse events were reported in 131 of the 532 patients who received remdesivir (24.6%) and in 163 of the 516 patients in the placebo group (31.6%).
This study also reported an association between remdesivir administration and both clinical improvement and a lack of progression to more invasive respiratory intervention in patients receiving non-invasive and invasive ventilation at randomization [435].
Largely on the results of this trial, the FDA reissued and expanded the EUA for remdesivir for the treatment of hospitalized COVID-19 patients ages twelve and older [432].
Additional clinical trials [439,446,447,448,449] are currently underway to evaluate the use of remdesivir to treat COVID-19 patients at both early and late stages of infection and in combination with other drugs (Figure 3).
As of October 22, 2020, remdesivir received FDA approval based on three clinical trials [450].
However, results suggesting no effect of remdesivir on survival were reported by the WHO Solidarity trial [424].
This large-scale, open-label trial enrolled 11,330 adult in-patients at 405 hospitals in 30 countries around the world [424].
Patients were randomized in equal proportions into four experimental conditions and a control condition, corresponding to four candidate treatments for COVID-19 and SOC, respectively; no placebo was administered.
The 2,750 patients in the remdesivir group were administered 200 mg intravenously on the first day and 100 mg on each subsequent day until day 10 and assessed for in-hospital death (primary endpoint), duration of hospitalization, and progression to mechanical ventilation.
There were also 2,708 control patients who would have been eligible and able to receive remdesivir were they not assigned to the control group.
A total of 604 patients among these two cohorts died during initial hospitalization, with 301 in the remdesivir group and 303 in the control group.
The rate ratio of death between these two groups was therefore not significant (0.95, p = 0.50), suggesting that the administration of remdesivir did not affect survival.
The two secondary analyses similarly did not find any effect of remdesivir.
Additionally, the authors compared data from their study with data from three other studies of remdesivir (including [435]) stratified by supplemental oxygen status.
A meta-analysis of the four studies yielded an overall rate ratio for death of 0.91 (p = 0.20).
These results thus do not support the previous findings that remdesivir reduced median recovery time and mortality risk in COVID-19 patients.
In response to the results of the Solidarity trial, Gilead, which manufactures remdesivir, released a statement pointing to the fact that the Solidarity trial was not placebo-controlled or double-blind and at the time of release, the statement had not been peer reviewed [451]; these sentiments have been echoed elsewhere [452].
Other critiques of this study have noted that antivirals are not typically targeted at patients with severe illness, and therefore remdesivir could be more beneficial for patients with mild rather than severe cases [426,453].
However, the publication associated with the trial sponsored by Gilead did purport an effect of remdesivir on patients with severe disease, identifying an 11 versus 18 day recovery period (rate ratio for recovery: 1.31, 95% CI 1.12 to 1.52) [435].
Additionally, a smaller analysis of 598 patients, of whom two-thirds were randomized to receive remdesivir for either 5 or 10 days, reported a small effect of treatment with remdesivir for five days relative to standard of care in patients with moderate COVID-19 [454].
These results suggest that remdesivir could improve outcomes for patients with moderate COVID-19, but that additional information would be needed to understand the effects of different durations of treatment.
Therefore, the Solidarity trial may point to limitations in the generalizability of other research on remdesivir, especially since the broad international nature of the Solidarity clinical trial, which included countries with a wide range of economic profiles and a variety of healthcare systems, provides a much-needed global perspective in a pandemic [426].
On the other hand, only 62% of patients in the Solidarity trial were randomized on the day of admission or one day afterwards [424], and concerns have been raised that differences in disease progression could influence the effectiveness of remdesivir [426].
Despite the findings of the Solidarity trial, remdesivir remains available for the treatment of COVID-19 in many places.
Remdesivir has also been investigated in combination with other drugs, such as baricitinib, which is an inhibitor of Janus kinase 1 and 2 [455]; the FDA has issued an EUA for the combination of remdesivir and baricitinib in adult and pediatric patients [456].
Follow-up studies are needed and, in many cases, are underway to further investigate remdesivir-related outcomes.
Similarly, the extent to which the remdesivir dosing regimen could influence outcomes continues to be under consideration.
A randomized, open-label trial compared the effect of remdesivir on 397 patients with severe COVID-19 over 5 versus 10 days [433,445], complementing the study that found that a 5-day course of remdesivir improved outcomes for patients with moderate COVID-19 but a 10-day course did not [454].
Patients in the two groups were administered 200 mg of remdesivir intravenously on the first day, followed by 100 mg on the subsequent four or nine days, respectively.
The two groups differed significantly in their clinical status, with patients assigned to the 10-day group having more severe illness.
This study also differed from most because it included not only adults, but also pediatric patients as young as 12 years old.
It reported no significant differences across several outcomes for patients receiving a 5-day or 10-day course, when correcting for baseline clinical status.
The data did suggest that the 10-day course might reduce mortality in the most severe patients at day 14, but the representation of this group in the study population was too low to justify any conclusions [445].
Thus, additional research is also required to determine whether the dosage and duration of remdesivir administration influences outcomes.
In summary, remdesivir is the first FDA approved anti-viral against SARS-CoV-2 as well as the first FDA approved COVID-19 treatment.
Early investigations of this drug established proof of principle that drugs targeting the virus can benefit COVID-19 patients.
It also shows proof of principle that SARS-CoV-2 can be targeted at the level of viral replication, since remdesivir targets the viral RNA polymerase at high potency.
Moreover, one of the most successful strategies for developing therapeutics for viral diseases is to target the viral replication machinery, which are typically virally encoded polymerases.
Small molecule drugs targeting viral polymerases are the backbones of treatments for other viral diseases including human immunodeficiency virus (HIV) and herpes.
Notably, the HIV and herpes polymerases are a reverse transcriptase and a DNA polymerase, respectively, whereas SARS-CoV-2 encodes an RdRP, so most of the commonly used polymerase inhibitors are not likely to be active against SARS-CoV-2.
In clinical use, polymerase inhibitors show short term benefits for HIV patients, but for long term benefits they must be part of combination regimens.
They are typically combined with protease inhibitors, integrase inhibitors, and even other polymerase inhibitors.
Additional clinical trials of remdesivir in different patient pools and in combination with other therapies will refine its use in the clinic.
5.4.1.2 Protease Inhibitors
Several studies showed that viral proteases play an important role in the life cycle of viruses, including coronaviruses, by modulating the cleavage of viral polyprotein precursors [457].
Several FDA-approved drugs target proteases, including lopinavir and ritonavir for HIV infection and simeprevir for hepatitis C virus infection.
In particular, serine protease inhibitors were suggested for the treatment of SARS and MERS viruses [458].
Recently, a study [70] suggested that camostat mesylate, an FDA-approved protease inhibitor could block the entry of SARS-CoV-2 into lung cells in vitro.
Thus far, investigation of possible protease inhibitors that could work against SARS-CoV-2 has been driven by computational predictions.
Computer-aided design allowed for the development of a Michael acceptor inhibitor, now known as N3, to target a protease critical to SARS-CoV-2 replication.
Discovery of the N3 mechanism arose from interest in the two polyproteins encoded by the SARS-CoV-2 replicase gene, pp1a and pp1ab, that are critical for viral replication and transcription [459].
These polyproteins must undergo proteolytic processing.
This processing is usually conducted by Mpro, a 33.8-kDa SARS-CoV-2 protease that is therefore fundamental to viral replication and transcription.
N3 was designed computationally [460] to bind in the substrate binding pocket of the Mpro protease of SARS-like coronaviruses [461], therefore inhibiting proteolytic processing.
Subsequently, the structure of N3-bound SARS-CoV-2 Mpro was solved [459], confirming the computational prediction.
N3 was tested in vitro on SARS-CoV-2-infected Vero cells, which belong to a line of cells established from the kidney epithelial cells of an African green monkey, and was found to inhibit SARS-CoV-2 [459].
Although N3 is a strong inhibitor of SARS-CoV-2 in vitro, its safety and efficacy still need to be tested in healthy volunteers and patients.
After the design and confirmation of N3 as a highly potent Michael acceptor inhibitor and the identification of Mpro’s structure [459,462], 10,000 compounds were screened for their in vitro anti-Mpro activity.
The six leads that were identified were ebselen, disulfiram, tideglusib, carmofur, and PX-12.
In vitro analysis revealed that ebselen had the strongest potency in reducing the viral load in SARS-CoV-2-infected Vero cells [459].
Ebselen is an organoselenium compound with anti-inflammatory and antioxidant properties [463].
It has been proposed as a possible treatment for conditions ranging from bipolar disorder to diabetes to heart disease [463], and a preliminary investigation of ebselen as a treatment for noise-induced hearing loss provided promising reports of its safety [464].
For COVID-19, the NSP5 in SARS-CoV-2 contains a cysteine at the active site of Mpro, and ebselen is able to inactivate the protease by bonding covalently with this cysteine to form a selenosulfide [463,465].
Interestingly there has been some argument that selenium deficiency may be associated with more severe COVID-19 outcomes [466,467,468], possibly indicating that its antioxidative properties are protective [465].
On the other hand, ebselen and the other compounds identified are likely to be promiscuous binders, which could diminish their therapeutic potential [459].
While there is clear computational and in vitro support for ebselen’s potential as a COVID-19 therapeutic, results from clinical trials are not yet available for this compound.
However, as of July 2020, phase II clinical trials commenced to assess the effects of SPI-1005, an investigational drug from Sound Pharmaceuticals that contains ebselen [469], on 60 adults presenting with each of moderate [470] and severe [471] COVID-19.
In summary, N3 is a computationally designed molecule that inhibits the viral transcription through inhibiting Mpro.
Ebselen is both a strong Mpro inhibitor and strong inhibitor of viral replication in vitro that was found to reduce SARS-CoV-2 viral load even more effectively than N3.
Ebselen is a promising compound since its safety has been demonstrated in other indications.
However, ebselen may be a false positive, since it is a promiscuous compound that can have many targets [472].
Therefore, the results of ongoing clinical trials are expected to help establish whether compounds with higher specificity are required.
5.4.2 Broad-Spectrum Pharmaceuticals
When a virus enters a host, the host becomes the virus’s environment.
Therefore, the state of the host can also influence the virus’s ability to replicate and spread.
Traditionally, viral targets have been favored for pharmaceutical interventions because altering host processes is likely to be less specific than targeting the virus directly [473].
On the other hand, targeting the host offers potential for a complementary strategy to antivirals that could broadly limit the ability of viruses to replicate [473].
As a result, therapeutic approaches that target host proteins have become an area of interest for SARS-CoV-2.
Viral entry receptors in particular have been identified as a potential target.
Entry of SARS-CoV-2 into the cell depends on binding to angiotensin-converting enzyme 2 (ACE2), which is catalyzed by the enzyme encoded by TMPRSS2[70].
In principle, drugs that reduce the expression of these proteins or sterically hinder viral interactions with them might reduce viral entry into cells.
Due to the urgent nature of the COVID-19 pandemic, many of the pharmaceutical agents that have been widely publicized as having possible therapeutic or prophylactic effects are broad-spectrum pharmaceuticals that pre-date the COVID-19 pandemic.
These treatments are not specifically targeted at the virus itself or at the host receptors it relies on, but rather induce broad shifts in host biology that are hypothesized to be potential inhibitors of the virus.
In most cases, interest in particular candidate medications arises because they are already available for other purposes.
However, the fact that the targets of these agents are non-specific means that the mechanism of action can appear to be relevant to COVID-19 without a therapeutic or prophylactic effect being observed in clinical trials.
This category of drugs has also received significant attention from the media and general public, often before rigorous testing has been able to determine their effectiveness against SARS-CoV-2.
5.4.2.1 ACE Inhibitors and Angiotensin II Receptor Blockers
Angiotensin-converting enzyme (ACE) inhibitors and angiotensin II receptor blockers (ARBs) are among today’s most commonly prescribed medications, often being used to control blood pressure [474,475].
In the United States, for example, they are prescribed well over 100,000,000 times annually [476].
Data from some animal models suggest that several, but not all, ACE inhibitors (ACEIs) and several ARBs increase ACE2 expression in the cells of some organs [477].
Clinical studies have not established whether plasma ACE2 expression is increased in humans treated with these medications [478].
While randomized clinical trials are ongoing, a variety of observational studies have examined the relationship between exposure to ACEIs or ARBs and outcomes in patients with COVID-19.
An observational study of the association of exposure to ACEIs or ARBs with outcomes in COVID-19 was retracted from the New England Journal of Medicine[479] due to concerns related to data availability [480].
Clinical trials are needed because the findings of the various observational studies bearing on this topic cannot be interpreted as indicating a protective effect of the drug [481,482].
Several clinical trials testing the effects of ACEIs or ARBs on COVID-19 outcomes are ongoing [483,484,485,486,487,488,489].
Two of these analyses [483,489] have reported no effect of continuing or discontinuing ARBs and ACEIs on patients admitted to the hospital for COVID-19.
The first, known as REPLACE COVID [490], was a randomized, open-label study that enrolled patients who were admitted to the hospital for COVID-19 and were taking an ACEI at the time of admission.
They enrolled 152 patients at 20 hospitals across seven countries and randomized them into two arms, continuation (n=75) and discontinuation (n=77).
The primary outcome evaluated was a global rank score that integrated several dimensions of illness.
The components of this global rank score, such as time to death and length of mechanical ventilation, were evaluated as secondary endpoints.
This analysis reported no differences between the two groups in the primary or any of the secondary outcomes.
Similarly, a second study [491] used a randomized, open-label design to examine the effects of continuing versus discontinuing ARBs and ACEIs on patients hospitalized for mild to moderate COVID-19 at 29 hospitals in Brazil.
This study enrolled 740 patients but had to exclude one trial site from all analyses due to the discovery of violations of Good Clinical Trial practice and data falsification.
After this exclusion, 659 patients remained, with 334 randomized to discontinuation and 325 to continuation.
In this study, the primary endpoint analyzed was the number of days that patients were alive and not hospitalized within 30 days of enrollment.
The secondary outcomes included death (including in-hospital death separately), number of days hospitalized, and specific clinical outcomes such as heart failure or stroke.
Once again, no significant differences were found between the two groups.
Initial studies of randomized interventions therefore suggest that ACEIs and ARBs are unlikely to affect COVID-19 outcomes.
These results are also consistent with findings from observational studies (summarized in [490]).
Additional information about ACE2, observational studies of ACEIs and ARBs in COVID-19, and clinical trials on this topic have been summarized [492].
Therefore, despite the promising potential mechanism, initial results have not provided support for ACEIs and ARBs as therapies for COVID-19.
5.4.2.2 Hydroxychloroquine and Chloroquine
CQ and hydroxychloroquine (HCQ) are lysosomotropic agents, meaning they are weak bases that can pass through the plasma membrane.
Both drugs increase cellular pH by accumulating in their protonated form inside lysosomes [493,494].
These drugs are used for the treatment and prophylaxis of malaria, as well as the treatment of lupus erythematosus and rheumatoid arthritis in adults [495].
This shift in pH inhibits the breakdown of proteins and peptides by the lysosomes during the process of proteolysis [494].
A number of mechanisms have been proposed through which these drugs could influence the immune response to pathogen challenge.
For example, CQ/HCQ can interfere with digestion of antigens within the lysosome and inhibit CD4 T-cell stimulation while promoting the stimulation of CD8 T-cells [494].
CQ/HCQ can also decrease the production of certain key cytokines involved in the immune response, including interleukin-6 (IL-6), and inhibit the stimulation of Toll-like receptors (TLR) and TLR signaling [494].
The drugs also have anti-inflammatory and photoprotective effects and may also affect rates of cell death, blood clotting, glucose tolerance, and cholesterol levels [494].
Interest in CQ and HCQ for treating COVID-19 was catalyzed by a mechanism observed in in vitro studies of both SARS-CoV-1 and SARS-CoV-2.
In one study, CQ inhibited viral entry of SARS-CoV-1 into Vero E6 cells, a cell line that was derived from Vero cells in 1968, through the elevation of endosomal pH and the terminal glycosylation of ACE2 [496].
Increased pH within the cell, as discussed above, inhibits proteolysis, and terminal glycosylation of ACE2 is thought to interfere with virus-receptor binding.
An in vitro study of SARS-CoV-2 infection of Vero cells found both HCQ and CQ to be effective in inhibiting viral replication, with HCQ being more potent [497].
Additionally, an early case study of three COVID-19 patients reported the presence of antiphospholipid antibodies in all three patients [119].
Antiphospholipid antibodies are central to the diagnosis of the antiphospholipid syndrome, a disorder that HCQ has often been used to treat [498,499,500].
Because the 90% effective concentration (EC90) of CQ in Vero E6 cells (6.90 μM) can be achieved in and tolerated by rheumatoid arthritis (RA) patients, it was hypothesized that it might also be possible to achieve the effective concentration in COVID-19 patients [501].
Additionally, clinical trials have reported HCQ to be effective in treating HIV [502] and chronic Hepatitis C [503].
Together, these studies triggered initial enthusiasm about the therapeutic potential for HCQ and CQ against COVID-19.
HCQ/CQ has been proposed both as a treatment for COVID-19 and a prophylaxis against SARS-CoV-2 exposure, and trials often investigated these drugs in combination with azithromycin (AZ) and/or zinc supplementation.
However, as more evidence has emerged, it has become clear that HCQ/CQ offer no benefits against SARS-CoV-2 or COVID-19.
5.4.2.2.1 Trials Assessing Therapeutic Administration of HCQ/CQ
The initial study evaluating HCQ as a treatment for COVID-19 patients was published on March 20, 2020 by Gautret et al. [504].
This non-randomized, non-blinded, non-placebo clinical trial compared HCQ to SOC in 42 hospitalized patients in southern France.
It reported that patients who received HCQ showed higher rates of virological clearance by nasopharyngeal swab on days 3-6 when compared to SOC.
This study also treated six patients with both HCQ + AZ and found this combination therapy to be more effective than HCQ alone.
However, the design and analyses used showed weaknesses that severely limit interpretability of results, including the small sample size and the lack of: randomization, blinding, placebo (no “placebo pill” given to SOC group), Intention-To-Treat analysis, correction for sequential multiple comparisons, and trial pre-registration.
Furthermore, the trial arms were entirely confounded by hospital and there were false negative outcome measurements (see [505]).
Two of these weaknesses are due to inappropriate data analysis and can therefore be corrected post hoc by recalculating the p-values (lack of Intention-To-Treat analysis and multiple comparisons).
However, all other weaknesses are fundamental design flaws and cannot be corrected for.
Thus, the conclusions cannot be generalized outside of the study.
The International Society of Antimicrobial Chemotherapy, the scientific organization that publishes the journal where the article appeared, subsequently announced that the article did not meet its expected standard for publications [506], although it has not been officially retracted.
Because of the preliminary data presented in this study, HCQ treatment was subsequently explored by other researchers.
About one week later, a follow-up case study reported that 11 consecutive patients were treated with HCQ + AZ using the same dosing regimen [507].
One patient died, two were transferred to the intensive care unit (ICU), and one developed a prolonged QT interval, leading to discontinuation of HCQ + AZ administration.
As in the Gautret et al. study, the outcome assessed was virological clearance at day 6 post-treatment, as measured from nasopharyngeal swabs.
Of the ten living patients on day 6, eight remained positive for SARS-CoV-2 RNA.
Like in the original study, interpretability was severely limited by the lack of a comparison group and the small sample size.
However, these results stand in contrast to the claims by Gautret et al. that all six patients treated with HCQ + AZ tested negative for SARS-CoV-2 RNA by day 6 post-treatment.
This case study illustrated the need for further investigation using robust study design to evaluate the efficacy of HCQ and/or CQ.
On April 10, 2020, a randomized, non-placebo trial of 62 COVID-19 patients at the Renmin Hospital of Wuhan University was released [508].
This study investigated whether HCQ decreased time to fever break or time to cough relief when compared to SOC [508].
This trial found HCQ decreased both average time to fever break and average time to cough relief, defined as mild or no cough.
While this study improved on some of the methodological flaws in Gautret et al. by randomizing patients, it also had several flaws in trial design and data analysis that prevent generalization of the results.
These weaknesses include the lack of placebo, lack of correction for multiple primary outcomes, inappropriate choice of outcomes, lack of sufficient detail to understand analysis, drastic disparities between pre-registration [509] and published protocol (including differences in the inclusion and exclusion criteria, the number of experimental groups, the number of patients enrolled, and the outcome analyzed), and small sample size.
The choice of outcomes may be inappropriate as both fevers and cough may break periodically without resolution of illness.
Additionally, for these outcomes, the authors reported that 23 of 62 patients did not have a fever and 25 of 62 patients did not have a cough at the start of the study, but the authors failed to describe how these patients were included in a study assessing time to fever break and time to cough relief.
It is important to note here that the authors claimed “neither the research performers nor the patients were aware of the treatment assignments.”
This blinding seems impossible in a non-placebo trial because at the very least, providers would know whether they were administering a medication or not, and this knowledge could lead to systematic differences in the administration of care.
Correction for multiple primary outcomes can be adjusted post hoc by recalculating p-values, but all of the other issues were design and statistical weaknesses that cannot be corrected for.
Additionally, the observation of drastic disparities between pre-registration and published protocol could indicate p-hacking [510].
The design limitations mean that the conclusions cannot be generalized outside of the study.
A second randomized trial, conducted by the Shanghai Public Health Clinical Center, analyzed whether HCQ increased rates of virological clearance at day 7 in respiratory pharyngeal swabs compared to SOC [511].
This trial was published in Chinese along with an abstract in English, and only the English abstract was read and interpreted for this review.
The trial found comparable outcomes in virological clearance rate, time to virological clearance, and time to body temperature normalization between the treatment and control groups.
The small sample size is one weakness, with only 30 patients enrolled and 15 in each arm.
This problem suggests the study is underpowered to detect potentially useful differences and precludes interpretation of results.
Additionally, because only the abstract could be read, other design and analysis issues could be present.
Thus, though these studies added randomization to their assessment of HCQ, their conclusions should be interpreted very cautiously.
These two studies assessed different outcomes and reached differing conclusions about the efficacy of HCQ for treating COVID-19; the designs of both studies, especially with respect to sample size, meant that no general conclusions can be made about the efficacy of the drug.
Several widely reported studies on HCQ also have issues with data integrity and/or provenance.
A Letter to the Editor published in BioScience Trends on March 16, 2020 claimed that numerous clinical trials have shown that HCQ is superior to control treatment in inhibiting the exacerbation of COVID-19 pneumonia [512].
This letter has been cited by numerous primary literature, review articles, and media alike [513,514].
However, the letter referred to 15 pre-registration identifiers from the Chinese Clinical Trial Registry.
When these identifiers are followed back to the registry, most trials claim they are not yet recruiting patients or are currently recruiting patients.
For all of these 15 identifiers, no data uploads or links to publications could be located on the pre-registrations.
At the very least, the lack of availability of the primary data means the claim that HCQ is efficacious against COVID-19 pneumonia cannot be verified.
Similarly, a recent multinational registry analysis [515] analyzed the efficacy of CQ and HCQ with and without a macrolide, which is a class of antibiotics that includes Azithromycin, for the treatment of COVID-19.
The study observed 96,032 patients split into a control and four treatment conditions (CQ with and without a macrolide; HCQ with and without a macrolide).
They concluded that treatment with CQ or HCQ was associated with increased risk of de novo ventricular arrhythmia during hospitalization.
However, this study has since been retracted by The Lancet due to an inability to validate the data used [516].
These studies demonstrate that increased skepticism in evaluation of the HCQ/CQ and COVID-19 literature may be warranted, possibly because of the significant attention HCQ and CQ have received as possible treatments for COVID-19 and the politicization of these drugs.
Despite the fact that the study suggesting that CQ/HCQ increased risk of ventricular arrhythmia in COVID-19 patients has now been retracted, previous studies have identified risks associated with HCQ/CQ.
A patient with systemic lupus erythematosus developed a prolonged QT interval that was likely exacerbated by use of HCQ in combination with renal failure [517].
A prolonged QT interval is associated with ventricular arrhythmia [518].
Furthermore, a separate study [519] investigated the safety associated with the use of HCQ with and without macrolides between 2000 and 2020.
The study involved 900,000 cases treated with HCQ and 300,000 cases treated with HCQ + AZ.
The results indicated that short-term use of HCQ was not associated with additional risk, but that HCQ + AZ was associated with an enhanced risk of cardiovascular complications (such as a 15% increased risk of chest pain, calibrated HR = 1.15, 95% CI, 1.05 to 1.26) and a two-fold increased 30-day risk of cardiovascular mortality (calibrated HR = 2.19; 95% CI, 1.22 to 3.94).
Therefore, whether studies utilize HCQ alone or HCQ in combination with a macrolide may be an important consideration in assessing risk.
As results from initial investigations of these drug combinations have emerged, concerns about the efficacy and risks of treating COVID-19 with HCQ and CQ have led to the removal of CQ/HCQ from SOC practices in several countries [520,521].
As of May 25, 2020, WHO had suspended administration of HCQ as part of the worldwide Solidarity Trial [522], and later the final results of this large-scale trial that compared 947 patients administered HCQ to 906 controls revealed no effect on the primary outcome, mortality during hospitalization (rate ratio: 1.19; p = 0.23)
Additional research has emerged largely identifying HCQ/CQ to be ineffective against COVID-19 while simultaneously revealing a number of significant side effects.
A randomized, open-label, non-placebo trial of 150 COVID-19 patients was conducted in parallel at 16 government-designated COVID-19 centers in China to assess the safety and efficacy of HCQ [523].
The trial compared treatment with HCQ in conjunction with SOC to SOC alone in 150 infected patients who were assigned randomly to the two groups (75 per group).
The primary endpoint of the study was the negative conversion rate of SARS-CoV-2 in 28 days, and the investigators found no difference in this parameter between the groups (estimated difference between SOC plus HCQ and SOC 4.1%; 95% CI, –10.3% to 18.5%).
The secondary endpoints were an amelioration of the symptoms of the disease such as axillary temperature ≤36.6°C, SpO2 >94% on room air, and disappearance of symptoms like shortness of breath, cough, and sore throat.
The median time to symptom alleviation was similar across different conditions (19 days in HCQ + SOC versus 21 days in SOC, p = 0.97).
Additionally, 30% of the patients receiving SOC+HCQ reported adverse outcomes compared to 8.8% of patients receiving only SOC, with the most common adverse outcome in the SOC+HCQ group being diarrhea (10% versus 0% in the SOC group, p = 0.004).
However, there are several factors that limit the interpretability of this study.
Most of the enrolled patients had mild-to-moderate symptoms (98%), and the average age was 46.
SOC in this study included the use of antivirals (Lopinavir-Ritonavir, Arbidol, Oseltamivir, Virazole, Entecavir, Ganciclovir, and Interferon alfa), which the authors note could influence the results.
Thus, they note that an ideal SOC would need to exclude the use of antivirals, but that ceasing antiviral treatment raised ethical concerns at the time that the study was conducted.
In this trial, the samples used to test for the presence of the SARS-CoV-2 virus were collected from the upper respiratory tract, and the authors indicated that the use of upper respiratory samples may have introduced false negatives (e.g., [97]).
Another limitation of the study that the authors acknowledge was that the HCQ treatment began, on average, at a 16-day delay from the symptom onset.
The fact that this study was open-label and lacked a placebo limits interpretation, and additional analysis is required to determine whether HCQ reduces inflammatory response.
Therefore, despite some potential areas of investigation identified in post hoc analysis, this study cannot be interpreted as providing support for HCQ as a therapeutic against COVID-19.
This study provided no support for HCQ against COVID-19, as there was no difference between the two groups in either negative seroconversion at 28 days or symptom alleviation, and in fact, more severe adverse outcomes were reported in the group receiving HCQ.
Additional evidence comes from a retrospective analysis [524] that examined data from 368 COVID-19 patients across all United States Veteran Health Administration medical centers.
The study retrospectively investigated the effect of the administration of HCQ (n=97), HCQ + AZ (n=113), and no HCQ (n=158) on 368 patients.
The primary outcomes assessed were death and the need for mechanical ventilation.
Standard supportive care was rendered to all patients.
Due to the low representation of women (N=17) in the available data, the analysis included only men, and the median age was 65 years.
The rate of death was 27.8% in the HCQ-only treatment group, 22.1% in the HCQ + AZ treatment group, and 14.1% in the no-HCQ group.
These data indicated a statistically significant elevation in the risk of death for the HCQ-only group compared to the no-HCQ group (adjusted HR: 2.61, p = 0.03), but not for the HCQ + AZ group compared to the no-HCQ group (adjusted HR: 1.14; p = 0.72).
Further, the risk of ventilation was similar across all three groups (adjusted HR: 1.43, p = 0.48 (HCQ) and 0.43, p = 0.09 (HCQ + AZ) compared to no HCQ).
The study thus showed evidence of an association between increased mortality and HCQ in this cohort of COVID-19 patients but no change in rates of mechanical ventilation among the treatment conditions.
The study had a few limitations: it was not randomized, and the baseline vital signs, laboratory tests, and prescription drug use were significantly different among the three groups.
All of these factors could potentially influence treatment outcome.
Furthermore, the authors acknowledge that the effect of the drugs might be different in females and pediatric subjects, since these subjects were not part of the study.
The reported result that HCQ + AZ is safer than HCQ contradicts the findings of the previous large-scale analysis of twenty years of records that found HCQ + AZ to be more frequently associated with cardiac arrhythmia than HCQ alone [519]; whether this discrepancy is caused by the pathology of COVID-19, is influenced by age or sex, or is a statistical artifact is not presently known.
Finally, findings from the RECOVERY trial were released on October 8, 2020.
This study used a randomized, open-label design to study the effects of HCQ compared to SOC at 176 hospitals in the United Kingdom [525].
This large study enrolled 11,197 hospitalized patients whose physicians believed it would not harm them to participate.
Patients were randomized into either the control group or one of the treatment arms, with twice as many patients enrolled in the control group as any treatment group.
Of the patients eligible to receive HCQ, 1,561 were randomized into the HCQ arm, and 3,155 were randomized into the control arm.
The demographics of the HCQ and control groups were similar in terms of average age (65 years), proportion female (approximately 38%), ethnic make-up (73% versus 76% white), and prevalence of pre-existing conditions (56% versus 57% overall).
In the HCQ arm of the study, patients received 800 mg at baseline and again after 6 hours, then 400 mg at 12 hours and every subsequent 12 hours.
The primary outcome analyzed was all-cause mortality, and patient vital statistics were reported by physicians upon discharge or death, or else at 28 days following HCQ administration if they remained hospitalized.
The secondary outcome assessed was the combined risk of progression to invasive mechanical ventilation or death within 28 days.
By the advice of an external data monitoring committee, the HCQ arm of the study was reviewed early, leading to it being closed due a lack of support for HCQ as a treatment for COVID-19.
The rates of COVID-19-related mortality reported in the RECOVERY trial did not differ between the control and HCQ arms (rate ratio, 1.09; 95% CI, 0.97 to 1.23; p = 0.15), but patients receiving HCQ were slightly more likely to die due to cardiac events (0.4 percentage points).
Patients who received HCQ also had a longer duration of hospitalization than patients receiving usual care, being less likely to be discharged alive within 28 days (rate ratio 0.90; 95% CI, 0.83 to 0.98), and were more likely to progress to mechanical ventilation or death (risk ratio 1.14; 95% CI, 1.03 to 1.27).
This large-scale study thus builds upon studies in the United States and China to suggest that HCQ is not an effective treatment, and in fact may negatively impact COVID-19 patients due to its side effects.
Therefore, though none of the studies have been blinded, examining them together makes it clear that the available evidence points to significant dangers associated with the administration of HCQ to hospitalized COVID-19 patients, without providing any support for its efficacy.
5.4.2.2.2 HCQ for the Treatment of Mild Cases
One additional possible therapeutic application of HCQ considered was the treatment of mild COVID-19 cases in otherwise healthy individuals.
This possibility was assessed in a randomized, open-label, multi-center analysis conducted in Catalonia (Spain) [526].
This analysis enrolled adults 18 and older who had been experiencing mild symptoms of COVID-19 for fewer than five days.
Participants were randomized into an HCQ arm (N=136) and a control arm (N=157), and those in the treatment arm were administered 800 mg of HCQ on the first day of treatment followed by 400 mg on each of the subsequent six days.
The primary outcome assessed was viral clearance at days 3 and 7 following the onset of treatment, and secondary outcomes were clinical progression and time to complete resolution of symptoms.
No significant differences between the two groups were found: the difference in viral load between the HCQ and control groups was 0.01 (95% CI, -0.28 to 0.29) at day 3 and -0.07 (95% CI -0.44 to 0.29) at day 7, the relative risk of hospitalization was 0.75 (95% CI, 0.32 to 1.77), and the difference in time to complete resolution of symptoms was -2 days (p = 0.38).
This study thus suggests that HCQ does not improve recovery from COVID-19, even in otherwise healthy adult patients with mild symptoms.
5.4.2.2.3 Prophylactic Administration of HCQ
An initial study of the possible prophylactic application of HCQ utilized a randomized, double-blind, placebo-controlled design to analyze the administration of HCQ prophylactically [527].
Asymptomatic adults in the United States and Canada who had been exposed to SARS-CoV-2 within the past four days were enrolled in an online study to evaluate whether administration of HCQ over five days influenced the probability of developing COVID-19 symptoms over a 14-day period.
Of the participants, 414 received HCQ and 407 received a placebo.
No significant difference in the rate of symptomatic illness was observed between the two groups (11.8% HCQ, 14.3% placebo, p = 0.35).
The HCQ condition was associated with side effects, with 40.1% of patients reporting side effects compared to 16.8% in the control group (p < 0.001).
However, likely due to the high enrollment of healthcare workers (66% of participants) and the well-known side effects associated with HCQ, a large number of participants were able to correctly identify whether they were receiving HCQ or a placebo (46.5% and 35.7%, respectively).
Furthermore, due to a lack of availability of diagnostic testing, only 20 of the 107 cases were confirmed with a PCR-based test to be positive for SARS-CoV-2.
The rest were categorized as “probable” or “possible” cases by a panel of four physicians who were blind to the treatment status.
One possible confounder is that a patient presenting one or more symptoms, which included diarrhea, was defined as a “possible” case, but diarrhea is also a common side effect of HCQ.
Additionally, four of the twenty PCR-confirmed cases did not develop symptoms until after the observation period had completed, suggesting that the 14-day trial period may not have been long enough or that some participants also encountered secondary exposure events.
Finally, in addition to the young age of the participants in this study, which ranged from 32 to 51, there were possible impediments to generalization introduced by the selection process, as 2,237 patients who were eligible but had already developed symptoms by day 4 were enrolled in a separate study.
It is therefore likely that asymptomatic cases were over-represented in this sample, which would not have been detected based on the diagnostic criteria used.
Therefore, while this study does represent the first effort to conduct a randomized, double-blind, placebo-controlled investigation of HCQ’s effect on COVID-19 prevention after SARS-CoV-2 exposure in a large sample, the lack of PCR tests and several other design flaws significantly impede interpretation of the results.
However, in line with the results from therapeutic studies, once again no evidence was found suggesting an effect of HCQ against COVID-19.
A second study [528] examined the effect of administering HCQ to healthcare workers as a pre-exposure prophylactic.
The primary outcome assessed was the conversion from SARS-CoV-2 negative to SARS-CoV-2 positive status over the 8 week study period.
This study was also randomized, double-blind, and placebo-controlled, and it sought to address some of the limitations of the first prophylactic study.
The goal was to enroll 200 healthcare workers, preferentially those working with COVID-19 patients, at two hospitals within the University of Pennsylvania hospital system in Philadelphia, PA.
Participants were randomized 1:1 to receive either 600 mg of HCQ daily or a placebo, and their SARS-CoV-2 infection status and antibody status were assessed using RT-PCR and serological testing, respectively, at baseline, 4 weeks, and 8 weeks following the beginning of the treatment period.
The statistical design of the study accounted for interim analyses at 50 and 100 participants in case efficacy or futility of HCQ for prophylaxis became clear earlier than completion of enrollment.
The 139 individuals enrolled comprised a study population that was fairly young (average age 33) and made of largely of people who were white, women, and without pre-existing conditions.
At the second interim analysis, more individuals in the treatment group than the control group had contracted COVID-19 (4 versus 3), causing the estimated z-score to fall below the pre-established threshold for futility.
As a result, the trial was terminated early, offering additional evidence against the use of HCQ for prophylaxis.
5.4.2.2.4 Summary of HCQ/CQ Research Findings
Early in vitro evidence indicated that HCQ could be an effective therapeutic against SARS-CoV-2 and COVID-19, leading to significant media attention and public interest in its potential as both a therapeutic and prophylactic.
Initially it was hypothesized that CQ/HCQ might be effective against SARS-CoV-2 in part because CQ and HCQ have both been found to inhibit the expression of CD154 in T-cells and to reduce TLR signaling that leads to the production of pro-inflammatory cytokines [529].
Clinical trials for COVID-19 have more often used HCQ rather than CQ because it offers the advantages of being cheaper and having fewer side effects than CQ.
However, research has not found support for a positive effect of HCQ on COVID-19 patients.
Multiple clinical studies have already been carried out to assess HCQ as a therapeutic agent for COVID-19, and many more are in progress.
To date, none of these studies have used randomized, double-blind, placebo-controlled designs with a large sample size, which would be the gold standard.
Despite the design limitations (which would be more likely to produce false positives than false negatives), initial optimism about HCQ has largely dissipated.
The most methodologically rigorous analysis of HCQ as a prophylactic [527] found no significant differences between the treatment and control groups, and the WHO’s global Solidarity trial similarly reported no effect of HCQ on mortality [424].
Thus, HCQ/CQ are not likely to be effective therapeutic or prophylactic agents against COVID-19.
Additionally, one study identified an increased risk of mortality in older men receiving HCQ, and administration of HCQ and HCQ + AZ did not decrease the use of mechanical ventilation in these patients [524].
HCQ use for COVID-19 could also lead to shortages for anti-malarial or anti-rheumatic use, where it has documented efficacy.
Despite significant early attention, these drugs appear to be ineffective against COVID-19.
Several countries have now removed CQ/HCQ from their SOC for COVID-19 due to the lack of evidence of efficacy and the frequency of adverse effects.
5.4.2.3 Dexamethasone
Dexamethasone (9α-fluoro-16α-methylprednisolone) is a synthetic corticosteroid that binds to glucocorticoid receptors [530,531].
It was first synthesized in the late 1950s as an anti-inflammatory and has been used to treat RA and other inflammatory conditions [532,533], including allergies and asthma [534].
Steroids such as dexamethasone are widely available and affordable, and they are often used to treat community-acquired pneumonia [535].
A clinical trial that began in 2012 recently reported that dexamethasone may improve outcomes for patients with ARDS [536].
However, a meta-analysis of a small amount of available data about dexamethasone as a treatment for SARS suggested that it may, in fact, be associated with patient harm [537]; these findings may have been biased by the fact that all of the studies examined were observational and a large number of inconclusive studies were not included [538].
Dexamethasone works as an anti-inflammatory agent by binding to glucocorticoid receptors with higher affinity than endogenous cortisol [539].
In order to understand how dexamethasone reduces inflammation, it is necessary to consider the stress response broadly.
In response to stress, corticotropin‐releasing hormone stimulates the release of neurotransmitters known as catecholamines, such as epinephrine, and steroid hormones known as glucocorticoids, such as cortisol [540,541].
While catecholamines are often associated with the fight-or-flight response, the specific role that glucocorticoids play is less clear, although they are thought to be important to restoring homeostasis [542].
Immune challenge is a stressor that is known to interact closely with the stress response.
The immune system can therefore interact with the central nervous system; for example, macrophages can both respond to and produce catecholamines [540].
Additionally, the production of both catecholamines and glucocorticoids is associated with inhibition of proinflammatory cytokines such as IL-6, IL-12, and tumor necrosis factor-α (TNF‐α) and the stimulation of anti-inflammatory cytokines such as IL-10, meaning that the stress response can regulate inflammatory immune activity [541].
Administration of dexamethasone has been found to correspond to dose-dependent inhibition of IL-12 production, but not to affect IL-10 [543]; the fact that this relationship could be disrupted by administration of a glucocorticoid-receptor antagonist suggests that it is regulated by the receptor itself [543].
Thus, the administration of dexamethasone for COVID-19 is likely to simulate the release of glucocorticoids endogenously during stress, resulting in binding of the synthetic steroid to the glucocorticoid receptor and the associated inhibition of the production of proinflammatory cytokines.
In this model, dexamethasone reduces inflammation by stimulating the biological mechanism that reduces inflammation following a threat such as immune challenge.
Immunosuppressive drugs such as steroids are typically contraindicated in the setting of infection [544], but because COVID-19 results in hyperinflammation that appears to contribute to mortality via lung damage, immunosuppression may be a helpful approach to treatment [161].
The decision of whether and/or when to counter hyperinflammation with immunosuppression in the setting of COVID-19 was an area of intense debate, as the risks of inhibiting antiviral immunity needed to be weighed against the beneficial anti-inflammatory effects [545].
As a result, guidelines early in the pandemic typically recommended avoiding treating COVID-19 patients with corticosteroids such as dexamethasone [537].
The application of dexamethasone for the treatment of COVID-19 was evaluated as part of the multi-site RECOVERY trial in the United Kingdom [546].
Over 6,000 hospitalized COVID-19 patients were assigned into the SOC or treatment (dexamethasone) arms of the trial with a 2:1 ratio.
At the time of randomization, some patients were ventilated (16%), others were on non-invasive oxygen (60%), and others were breathing independently (24%).
Patients in the treatment arm were administered dexamethasone either orally or intravenously at 6 mg per day for up to 10 days.
The primary end-point was the patient’s status at 28-days post-randomization (mortality, discharge, or continued hospitalization), and secondary outcomes analyzed included the progression to invasive mechanical ventilation over the same period.
The 28-day mortality rate was found to be lower in the treatment group than in the SOC group (21.6% vs 24.6%, p < 0.001).
However, this finding was driven by differences in mortality among patients who were receiving mechanical ventilation or supplementary oxygen at the start of the study.
The report indicated that dexamethasone reduced 28-day mortality relative to SOC in patients who were ventilated (29.3% versus 41.4%) and among those who were receiving oxygen supplementation (23.3% versus 26.2%) at randomization, but not in patients who were breathing independently (17.8% versus 14.0%).
One possible confounder is that patients receiving mechanical ventilation tended to be younger than patients who were not receiving respiratory support (by 10 years on average) and to have had symptoms for a longer period.
However, adjusting for age did not change the conclusions, although the duration of symptoms was found to be significantly associated with the effect of dexamethasone administration.
These findings also suggested that dexamethasone may have reduced progression to mechanical ventilation, especially among patients who were receiving oxygen support at randomization.
Thus, this large, randomized, and multi-site, albeit not placebo-controlled, study suggests that administration of dexamethasone to patients who are unable to breathe independently may significantly improve survival outcomes.
Additionally, dexamethasone is a widely available and affordable medication, raising the hope that it could be made available to COVID-19 patients globally.
The results of the RECOVERY trial’s analysis of dexamethasone suggest that this therapeutic is effective primarily in patients who had been experiencing symptoms for at least seven days and patients who were not breathing independently [547].
A meta-analysis that evaluated the results of the RECOVERY trial alongside trials of other corticosteroids, such as hydrocortisone, similarly concluded that corticosteroids may be beneficial to patients with severe COVID-19 who are receiving oxygen supplementation [548].
Thus, it seems likely that dexamethasone is useful for treating inflammation associated with immunopathy or cytokine release syndrome (CRS), which is a condition caused by detrimental overactivation of the immune system [1].
In fact, corticosteroids such as dexamethasone are sometimes used to treat CRS [549].
It is not surprising that administration of an immunosuppressant would be most beneficial when the immune system was dysregulated towards inflammation.
However, it is also unsurprising that care must be taken in administering an immunosuppressant to patients fighting a viral infection.
In particular, the concern has been raised that treatment with dexamethasone might increase patient susceptibility to concurrent (e.g., nosocomial) infections [550].
Additionally, the drug could potentially slow viral clearance and inhibit patients’ ability to develop antibodies to SARS-CoV-2 [537,550], with the lack of data about viral clearance being put forward as a major limitation of the RECOVERY trial [551].
Furthermore, dexamethasone has been associated with side effects that include psychosis, glucocorticoid-induced diabetes, and avascular necrosis [537], and the RECOVERY trial did not report outcomes with enough detail to be able to determine whether they observed similar complications.
The effects of dexamethasone have also been found to differ among populations, especially in high-income versus middle- or low-income countries [552].
However, since the RECOVERY trial’s results were released, strategies have been proposed for administering dexamethasone alongside more targeted treatments to minimize the likelihood of negative side effects [550].
Given the available evidence, dexamethasone is currently the most promising treatment for severe COVID-19.
5.5 Biologics
Biologics are produced from components of living organisms or viruses.
They include treatments such as humanized monoclonal antibodies, tocilizumab (TCZ), and neutralizing antibodies (nAbs), and can also include prophylactics such as vaccines [401].
Historically produced from animal tissue, biologics have become increasingly feasible to produce as recombinant DNA technologies have advanced [553].
Often, they are glycoproteins or peptides [554], but whole viruses can also be used therapeutically or prophylactically, not only for vaccines but also as vectors for gene therapy or therapeutic proteins or for oncolytic virotherapy [555].
They are typically catabolized by the body to their amino acid components [554].
There are many differences on the development side between biologics and synthesized pharmaceuticals, such as small molecule drugs.
Biologics are typically orders of magnitude larger than small molecule drugs, and their physiochemical properties are often much less understood [554].
They are often heat sensitive, and their toxicity can vary, as it is not directly associated with the primary effects of the drug [554].
However, this class includes some extremely significant medical breakthroughs, including insulin for the management of diabetes and the smallpox vaccine.
As a result, biologics are another possible avenue through which the pharmacological management of SARS-CoV-2 infection can be approached.
5.5.1 Tocilizumab
TCZ is a receptor antibody that was developed to manage chronic inflammation caused by the continuous synthesis of the cytokine IL-6 [556].
IL-6 is a pro-inflammatory cytokine belonging to the interleukin family, which is comprised by immune system regulators that are primarily responsible for immune cell differentiation.
Often used to treat conditions such as RA [556], TCZ has become a pharmaceutical of interest for the treatment of COVID-19 because of the role IL-6 plays in this disease.
It has also been approved to treat CRS caused by CAR-T treatments [557].
While secretion of IL-6 can be associated with chronic conditions, it is a key player in the innate immune response and is secreted by macrophages in response to the detection of pathogen-associated molecular patterns and damage-associated molecular patterns [556].
An analysis of 191 in-patients at two Wuhan hospitals revealed that blood concentrations of IL-6 differed between patients who did and did not recover from COVID-19.
Patients who ultimately died had higher IL-6 levels at admission than those who recovered [38].
Additionally, IL-6 levels remained higher throughout the course of hospitalization in the patients who ultimately died [38].
This finding provided some early evidence that COVID-19 deaths may be induced by the hyperactive immune response, often referred to as CRS or cytokine storm syndrome (CSS), as IL-6 plays a key role in this response [148].
In this context, the observation of elevated IL-6 in patients who died may reflect an over-production of proinflammatory interleukins, suggesting that TCZ could potentially palliate some of the most severe symptoms of COVID-19 associated with increased cytokine production.
Human IL-6 is a 26-kDa glycoprotein that consists of 184 amino acids and contains two potential N-glycosylation sites and four cysteine residues.
It binds to a type I cytokine receptor (IL-6Rα or glycoprotein 80) that exists in both membrane-bound (IL-6Rα) and soluble (sIL-6Rα) forms [558].
It is not the binding of IL-6 to the receptor that initiates pro- and/or anti-inflammatory signaling, but rather the binding of the complex to another subunit, known as IL-6Rβ or glycoprotein 130 (gp130) [558,559].
Unlike membrane-bound IL-6Rα, which is only found on hepatocytes and some types of leukocytes, gp130 is found on most cells [560].
When IL-6 binds to sIL-6Rα, the complex can then bind to a gp130 protein on any cell [560].
The binding of IL-6 to IL-6Rα is termed classical signaling, while its binding to sIL-6Rα is termed trans-signaling [560,561,562].
These two signaling processes are thought to play different roles in health and illness.
For example, trans-signaling may play a role in the proliferation of mucosal T-helper TH2 cells associated with asthma, while an earlier step in this proliferation process may be regulated by classical signaling [560].
Similarly, IL-6 is known to play a role in Crohn’s Disease via trans-, but not classical, signaling [560].
Both classical and trans-signaling can occur through three independent pathways: the Janus-activated kinase-STAT3 pathway, the Ras/Mitogen-Activated Protein Kinases pathway and the Phosphoinositol-3 Kinase/Akt pathway [558].
These signaling pathways are involved in a variety of different functions, including cell type differentiation, immunoglobulin synthesis, and cellular survival signaling pathways, respectively [558].
The ultimate result of the IL-6 cascade is to direct transcriptional activity of various promoters of pro-inflammatory cytokines, such as IL-1, TFN, and even IL-6 itself, through the activity of NF-κB [558].
IL-6 synthesis is tightly regulated both transcriptionally and post-transcriptionally, and it has been shown that viral proteins can enhance transcription of the IL-6 gene by strengthening the DNA-binding activity between several transcription factors and IL-6 gene-cis-regulatory elements [563].
Therefore, drugs inhibiting the binding of IL-6 to IL-6Rα or sIL-6Rα are of interest for combating the hyperactive inflammatory response characteristic of CRS/CSS.
TCZ is a humanized monoclonal antibody that binds both to the insoluble and soluble receptor of IL-6, providing de facto inhibition of the IL-6 immune cascade.
Tocilizumab is being administered either as an intervention or as concomitant medication in 77 interventional COVID-19 clinical trials (Figure 3).
No randomized, placebo-controlled studies of TCZ have currently released results.
Therefore, no conclusions can be drawn about its efficacy for the treatment of COVID-19.
However, early interest in TCZ as a possible treatment for COVID-19 emerged from a very small retrospective study in China that examined 20 patients with severe symptoms in early February 2020 and reported rapid improvement in symptoms following treatment with TCZ [564].
Subsequently, a number of retrospective studies have been conducted in several countries.
Many studies use a retrospective, observational design, where they compare outcomes for COVID-19 patients who received TCZ to those who did not over a set period of time.
For example, one of the largest retrospective, observational analyses released to date [565], consisting of 1,351 patients admitted to several care centers in Italy, compared the rates at which patients who received TCZ died or progressed to invasive medical ventilation over a 14-day period compared to patients receiving only SOC.
Under this definition, SOC could include other drugs such as HCQ, azithromycin, lopinavir-ritonavir or darunavir-cobicistat, or heparin.
While this study was not randomized, a subset of patients who were eligible to receive TCZ were unable to obtain it due to shortages; however, these groups were not directly compared in the analysis.
After adjusting for variables such as age, sex, and SOFA (sequential organ failure assessment) score, they found that patients treated with TCZ were less likely to progress to invasive medical ventilation and/or death (adjusted HR = 0.61, CI 0.40-0.92, p = 0.020); analysis of death and ventilation separately suggests that this effect may have been driven by differences in the death rate (20% of control versus 7% of TCZ-treated patients).
The study reported particular benefits for patients whose PaO2/FiO2 ratio, also known as the Horowitz Index for Lung Function, fell below a 150 mm Hg threshold.
They found no differences between groups administered subcutaneous versus intravenous TCZ.
Another retrospective observational analysis of interest examined the charts of patients at a hospital in Connecticut, USA where 64% of all 239 COVID-19 patients in the study period were administered TCZ based on assignment by a standardized algorithm [566].
They found that TCZ administration was associated with more similar rates of survivorship in patients with severe versus nonsevere COVID-19 at intake, defined based on the amount of supplemental oxygen needed.
They therefore proposed that their algorithm was able to identify patients presenting with or likely to develop CRS as good candidates for TCZ.
This study also reported higher survivorship in Black and Hispanic patients compared to white patients when adjusted for age.
The major limitation with interpretation for these studies is that there may be clinical characteristics that influenced medical practitioners decisions to administer TCZ to some patients and not others.
One interesting example therefore comes from an analysis of patients at a single hospital in Brescia, Italy, where TCZ was not available for a period of time [567].
This study compared COVID-19 patients admitted to the hospital before and after March 13, 2020, when the hospital received TCZ.
Therefore, patients who would have been eligible for TCZ prior to this arbitrary date did not receive it as treatment, making this retrospective analysis something of a natural experiment.
Despite this design, demographic factors did not appear to be consistent between the two groups, and the average age of the control group was older than the TCZ group.
The control group also had a higher percentage of males and a higher incidence of comorbidities such as diabetes and heart disease.
All the same, the multivariate HR, which adjusted for these clinical and demographic factors, found a significant difference between survival in the two groups (HR=0.035, CI=0.004-0.347, p = 0.004).
The study reported improvement of survival outcomes after the addition of TCZ to the SOC regime, with 11 of 23 patients (47.8%) admitted prior to March 13th dying compared to 2 of 62 (3.2%) admitted afterwards (HR=0.035; 95% CI, 0.004 to 0.347; p = 0.004).
They also reported a reduced progression to mechanical ventilation in the TCZ group.
However, this study also holds a significant limitation: the time delay between the two groups means that knowledge about how to treat the disease likely improved over this timeframe as well.
All the same, the results of these observational retrospective studies provide support for TCZ as a pharmaceutical of interest for follow-up in clinical trials.
Other retrospective analyses have utilized a case-control design to match pairs of patients with similar baseline characteristics, only one of whom received TCZ for COVID-19.
In one such study, TCZ was significantly associated with a reduced risk of progression to ICU admission or death [568].
This study examined only 20 patients treated with TCZ (all but one of the patients treated with TCZ in the hospital during the study period) and compared them to 25 patients receiving SOC.
For the combined primary endpoint of death and/or ICU admission, only 25% of patients receiving TCZ progressed to an endpoint compared to 72% in the SOC group (p = 0.002, presumably based on a chi-square test based on the information provided in the text).
When the two endpoints were examined separately, progression to invasive medical ventilation remained significant (32% SOC compared to 0% TCZ, p = 0.006) but not for mortality (48% SOC compared to 25% TCZ, p = 0.066).
In contrast, a study that compared 96 patients treated with TCZ to 97 patients treated with SOC only in New York City found that differences in mortality did not differ between the two groups, but that this difference did become significant when intubated patients were excluded from the analysis [569].
Taken together, these findings suggest that future clinical trials of TCZ may want to include intubation as an endpoint.
However, these studies should be approached with caution, not only because of the small number of patients enrolled and the retrospective design, but also because they performed a large number of statistical tests and did not account for multiple hypothesis testing.
In general, caution must be exercised when interpreting subgroup analyses after a primary combined endpoint analysis.
These last findings highlight the need to search for a balance between impairing a harmful immune response, such as the one generated during CRS/CSS, and preventing the worsening of the clinical picture of the patients by potential new viral infections.
Though data about TCZ for COVID-19 is still only just emerging, some meta-analyses and systematic reviews have investigated the available data.
One meta-analysis [570] evaluated 19 studies published or released as preprints prior to July 1, 2020 and found that the overall trends were supportive of the frequent conclusion that TCZ does improve survivorship, with a significant HR of 0.41 (p < 0.001).
This trend improved when they excluded studies that administered a steroid alongside TCZ, with a significant HR of 0.04 (p < 0.001).
They also found some evidence for reduced invasive ventilation or ICU admission, but only when excluding all studies except a small number whose estimates were adjusted for the possible bias introduced by the challenges of stringency during the enrollment process.
A systematic analysis of sixteen case-control studies of TCZ estimated an odds ratio of mortality of 0.453 (95% CI 0.376–0.547, p < 0.001), suggesting possible benefits associated with TCZ treatment [571].
Although these estimates are similar, it is important to note that they are drawing from the same literature and are therefore likely to be affected by the same potential biases in publication.
A different systematic review of studies investigating TCZ treatment for COVID-19 analyzed 31 studies that had been published or released as pre-prints and reported that none carried a low risk of bias [572].
Therefore, the present evidence is not likely to be sufficient for conclusions about the efficacy of TCZ.
On February 11, 2021, a preprint describing the first randomized control trial of TCZ was released as part of the RECOVERY trial [573].
Of the 21,550 patients enrolled in the RECOVERY trial at the time, 4,116 adults hospitalized with COVID-19 across the 131 sites in the United Kingdom were assigned to the arm of the trial evaluating the effect of TCZ.
Among them, 2,022 were randomized to receive TCZ and 2,094 were randomized to SOC, with 79% of patients in each group available for analysis at the time that the initial report was released.
The primary outcome measured was 28-day mortality, and TCZ was found to reduce 28-day mortality from 33% of patients receiving SOC alone to 29% of those receiving TCZ, corresponding to a rate ratio of 0.86 (95% CI 0.77-0.96; p = 0.007).
TCZ was also significantly associated with the probability of hospital discharge within 28 days for living patients, which was 47% in the SOC group and 54% in the TCZ group (rate ratio 1.22, 95% CI 1.12-1.34, p < 0.0001).
A potential statistical interaction between TCZ and corticosteroids was observed, with the combination providing greater mortality benefits than TCZ alone, but the authors note that caution is advisable in light of the number of statistical tests conducted.
Combining the RECOVERY trial data with data from seven smaller randomized control trials indicates that TCZ is associated with a 13% reduction in 28-day mortality (rate ratio 0.87, 95% CI 0.79-0.96, p = 0·005) [573].
While this initial report did not include the full results expected from the RECOVERY trial, this large-scale, randomized controlled trial therefore offers strong evidence that TCZ may offer benefits for COVID-19 patients, even at this initial stage of analysis.
There are possible risks associated with the administration of TCZ for COVID-19.
TCZ has been used for over a decade to treat RA [574], and a recent study found the drug to be safe for pregnant and breastfeeding women [575].
However, TCZ may increase the risk of developing infections [574], and RA patients with chronic hepatitis B infections had a high risk of hepatitis B virus reactivation when TCZ was administered in combination with other RA drugs [576].
As a result, TCZ is contraindicated in patients with active infections such as tuberculosis [577].
Previous studies have investigated, with varying results, a possible increased risk of infection in RA patients administered TCZ [578,579], although another study reported that the incidence rate of infections was higher in clinical practice RA patients treated with TCZ than in the rates reported by clinical trials [580].
In the investigation of 544 Italian COVID-19 patients, the group treated with TCZ was found to be more likely to develop secondary infections, with 24% compared to 4% in the control group (p < 0.0001) [565].
Reactivation of hepatitis B and herpes simplex virus 1 was also reported in a small number of patients in this study, all of whom were receiving TCZ.
A July 2020 case report described negative outcomes of two COVID-19 patients after receiving TCZ, including one death; however, both patients were intubated and had entered septic shock prior to receiving TCZ [581], likely indicating a severe level of cytokine production.
Additionally, D-dimer and sIL2R levels were reported by one study to increase in patients treated with TCZ, which raised concerns because of the potential association between elevated D-dimer levels and thrombosis and between sIL2R and diseases where T-cell regulation is compromised [566].
An increased risk of bacterial infection was also identified in a systematic review of the literature, based on the unadjusted estimates reported [570].
In the RECOVERY trial, however, only three out of 2,022 participants in the group receiving TCZ developed adverse reactions determined to be associated with the intervention, and no excess deaths were reported [573].
TCZ administration to COVID-19 patients is not without risks and may introduce additional risk of developing secondary infections; however, while caution may be prudent when treating patients who have latent viral infections, the results of the RECOVERY trial indicate that adverse reactions to TCZ are very rare among COVID-19 patients broadly.
In summary, approximately 33% of hospitalized COVID-19 patients develop ARDS [582], which is caused by an excessive early response of the immune system which can be a component of CRS/CSS [566,577].
This overwhelming inflammation is triggered by IL-6.
TCZ is an inhibitor of IL-6 and therefore may neutralize the inflammatory pathway that leads to the cytokine storm.
While the mechanism suggests TCZ could be beneficial for the treatment of COVID-19 patients experiencing excessive immune activity, no randomized controlled trials are available assessing its effect.
However, small initial studies have found preliminary indications that TCZ may reduce progression to invasive medical ventilation and/or death.
It should be noted that SOC varied widely across retrospective studies, with one study administering HCQ, lopinavir-ritonavir, antibiotics, and/or heparin as part of SOC.
Interest in TCZ as a treatment for COVID-19 was supported by two meta-analyses [570,583], but a third meta-analysis found that all of the available literature carries a risk of bias, with even the largest available TCZ studies to date carrying a moderate risk of bias under the ROBINS-I criteria [572].
Additionally, different studies used different dosages, number of doses, and methods of administration.
Ongoing research may be needed to optimize administration of TCZ [584], although similar results were reported by one study for intravenous and subcutaneous administration [565].
Clinical trials that are in progress are likely to provide additional insight into the effectiveness of this drug for the treatment of COVID-19 along with how it should be administered.
5.5.2 Monoclonal Neutralizing Antibodies
Monoclonal antibodies have revolutionized the way we treat human diseases.
They have become some of the best-selling drugs in the pharmaceutical market in recent years [585].
There are currently 79 FDA approved mAbs on the market, including antibodies for viral infections (e.g. Ibalizumab for HIV and Palivizumab for RSV) [585,586].
Virus-specific neutralizing antibodies commonly target viral surface glycoproteins or host structures, thereby inhibiting viral entry through receptor binding interference [587,588].
This is predicted to reduce the viral load, mitigate disease, and reduce overall hospitalization.
While polyclonal antibodies from convalescent plasma can be used as a treatment for COVID-19, this section focuses on current efforts in developing monoclonal neutralizing antibodies (nAbs) against SARS-CoV-2 (excellent reviews regarding convalescent plasma therapy can be found here [589,590]).
Specifically, we focus on monoclonal antibodies that have recently been granted emergency use authorization and discuss the challenges in the successful development of monoclonal neutralizing antibodies.
During the first SARS epidemic in 2002, nAbs were found in SARS-CoV-1-infected patients [591,592].
Several studies following up on these findings identified various S-glycoprotein epitopes as the major targets of nAbs against SARS-CoV-1 [593].
Coronaviruses use trimeric spike (S) glycoproteins on their surface to bind to the host cell, allowing for cell entry [67,70].
Each S glycoprotein protomer is comprised of an S1 domain, also called the RBD, and an S2 domain.
The S1 domain binds to the host cell while the S2 domain facilitates the fusion between the viral envelope and host cell membranes [593].
The genomic identity between the RBD of SARS-CoV-1 and SARS-CoV-2 is around 74% [594].
Due to this high degree of similarity, preexisting antibodies against SARS-CoV-1 were initially considered candidates for neutralizing activity against SARS-CoV-2.
While some antibodies developed against the SARS-CoV-1 spike protein showed cross-neutralization activity with SARS-CoV-2 [595,596], others failed to bind to SARS-CoV-2 spike protein at relevant concentrations [54].
Cross-neutralizing activities were dependent on whether the epitope recognized by the antibodies were conserved between SARS-CoV-1 and SARS-CoV-2 [595].
The first human monoclonal neutralizing antibody specifically against the SARS-CoV-2 S glycoprotein was developed using hybridoma technology [597], where antibody-producing B-cells developed by mice are inserted into myeloma cells to produce a hybrid cell line (the hybridoma) that is grown in culture.
The 47D11 antibody clone was able to cross-neutralize SARS-CoV-1 and SARS-CoV-2.
This antibody (now ABVV-47D11) has recently entered clinical trials in collaboration with AbbVie.
Since then, an extensive monoclonal neutralizing antibody pipeline has been developed to combat the ongoing pandemic, with over 50 different antibodies in clinical trials [598] and two treatments recently receiving emergency use authorization by the FDA.
5.5.2.1 Bamlanivimab (LY-CoV555) and Etesevimab (LY-CoV016)
Bamlanivimab is a human monoclonal antibody that was derived from convalescent plasma donated by recovered COVID-19 patient, evaluated in research by the National Institute of Allergy and Infectious Diseases (NIAID), and subsequently developed by AbCellera and Eli Lilly.
The neutralizing activity of bamlanivimab was initially demonstrated in vivo using a nonhuman primate model [599].
In these studies, prophylactic Ly-CoV555 infusions protected rhesus macaques from SARS-CoV-2 infection.
Based on the positive preclinical data, Eli Lilly initiated the first human clinical trial for a monoclonal antibody against SARS-CoV-2.
The phase 1 trial, which was conducted in hospitalized COVID-19 patients, was completed in August 2020 [600].
Estesevimab (LY-CoV016 or JS-016) is also a monoclonal neutralizing antibody against the spike protein of SARS-CoV-2.
It was initially developed by Junshi Biosciences and later licensed and developed through Eli Lilly.
A phase 1 clinical trial to assess the safety of etesevimab was completed in October 2020 [601].
Etesevimab was shown to bind a different epitope on the spike protein than bamlanivimab, suggesting that the two antibodies used as a combination therapy would further enhance their clinical use compared to a monotherapy [602].
To assess the efficacy and safety of bamlanivimab alone or in combination with etesevimab for the treatment of COVID-19, a phase 2/3 trial (BLAZE-1) [603] was initiated.
The interim analysis of the phase 2 portion suggested that bamlanivimab alone was able to reduce accelerate the reduction in viral load [604].
However, more recent data suggests that only the bamlanivimab/etesevimab combination therapy is able to reduce viral load in COVID-19 patients [602].
Based on this data, the combination therapy received emergency use authorization for COVID-19 from the FDA in February of 2021.
5.5.2.2 Casirivimab and Imdevimab (REGN-COV2)
Casirivimab (REGN10933) and imdevimab (REGN10987) are two monoclonal antibodies against the SARS-CoV-2 spike protein.
They were both developed by Regeneron in a parallel high-throughput screen to identify neutralizing antibodies from either humanized mice or patient-derived convalescent plasma [605].
In these efforts, multiple antibodies were characterized for their ability to bind and neutralize the SARS-CoV-2 spike protein.
The authors hypothesized that an antibody cocktail, rather than each individual antibody, could increase the therapeutic efficacy while minimizing the risk for virus escape.
Therefore, the authors tested pairs of individual antibodies for their ability to simultaneously bind the RBD of the spike protein.
Based on this data, casirivimab and imdevimab were identified as the lead antibody pair, resulting in the initiation of two clinical trials [606,607].
Data from this phase 1-3 trial published in the New England Journal of Medicine shows that the REGN-COV2 antibody cocktail reduced viral load, particularly in patients with high viral load or whose endogenous immune response had not yet been initiated [608].
However, in patients who already initiated an immune response, exogenous addition of REGN-COV2 did not improve the endogenous immune response.
Both doses were well tolerated with no serious events related to the antibody cocktail.
Based on this data, the FDA granted emergency use authorization for REGN-COV2 in patients with mild to moderate COVID-19 who are at risk to develop severe disease.
Ongoing efforts are trying to evaluate the efficacy of REGN-COV2 to improve clinical outcomes in hospitalized patients [606].
5.5.2.3 Viral Resistance to Neutralizing Antibodies
With the ongoing global spread of new SARS-CoV-2 variants, there is a growing concern that mutations in SARS-CoV-2 spike protein could escape antibody neutralization, thereby reducing the efficacy of monoclonal antibody therapeutics and vaccines.
A comprehensive mutagenesis screen recently identified several amino acid substitutions in the SARS-CoV-2 spike protein that can prevent antibody neutralization [609].
While some mutations result in resistance to only one antibody, others confer broad resistance to multiple monoclonal antibodies as well as polyclonal human sera, suggesting that some amino acids are “hotspots” for antibody resistance.
However, it was not investigated whether the resistant mutations identified result in a fitness advantage.
Accordingly, an impact on neutralizing efficiency has been reported for the emerging UK (B.1.1.7) and South Africa (B.1.351) variants [610,611,612].
While the reported impact on antibody neutralization needs to be confirmed in vivo, it suggests that some adjustments to therapeutic antibody treatments may be necessary to maintain the efficacy that was reported in previous clinical trials.
Antibody cocktails such as REGN-COV2, CT-P59, and AZD7442 have been developed to overcome the risk for attenuation of neutralizing activity of a single monoclonal antibody.
These cocktails consist of antibodies that recognize different epitopes on the spike protein, decreasing the likelihood that a single amino acid change can cause resistance to all antibodies in the cocktail.
However, neutralizing resistance can emerge even against an antibody cocktail if the individual antibodies target subdominant epitopes [611].
Another strategy is to develop broadly neutralizing antibodies that target structures that are highly conserved, as these are less likely to mutate [613,614] or to target epitopes that are insensitive to mutations [615].
One such antibody (ADG-2) has recently been reported [616].
This antibody targets a highly conserved epitope that overlaps the hACE2 binding site of all clade 1 sarbecoviruses.
Prophylactic administration of ADG-2 in an immunocompetent mouse model of COVID-19 resulted in protection against viral replication in the lungs and respiratory burden.
Since the epitope targeted by ADG-2 represents an Achilles’ heel for clade 1 sarbecoviruses, this antibody might be a promising candidate against all circulating variants as well as emerging SARS-related coronaviruses.
In just under a year since the structure of the SARS-CoV-2 spike protein was first published, an impressive pipeline of monoclonal antibodies targeting SARS-CoV-2 has entered clinical trials, with hundreds more candidates in preclinical stages.
Technological advances in antibody drug design as well as in structural biology massively accelerated the discovery of novel antibody candidates and the mechanisms by which they interact with the target structure.
One of the biggest challenges remains identifying antibodies that not only bind to their target, but also prove to be beneficial for disease management.
Thus far, two antibody cocktails (REGN-COV2 and LY-CoV555/LY-COV016) have been granted emergency use authorization by the FDA.
However, their current use is limited to people with mild to moderate disease that are not hospitalized.
Therefore, it has yet to be determined whether monoclonal antibodies can be used as a successful treatment option for severe COVID patients.
5.5.3 Interferons
IFNs are a family of cytokines critical to activating the innate immune response against viral infections.
Interferons are classified into three categories based on their receptor specificity: types I, II and III [148].
Specifically, IFNs I (IFN-𝛼 and 𝛽) and II (IFN-𝛾) induce the expression of antiviral proteins [617].
Among these IFNs, IFN-𝛽 has already been found to strongly inhibit the replication of other coronaviruses, such as SARS-CoV-1, in cell culture, while IFN-𝛼 and 𝛾 were shown to be less effective in this context [617].
There is evidence that patients with higher susceptibility to ARDS indeed show deficiency in IFN-𝛽.
For instance, infection with other coronaviruses impairs IFN-𝛽 expression and synthesis, allowing the virus to escape the innate immune response [618].
On March 18 2020, Synairgen plc received approval to start a phase II trial for SNG001, an IFN-𝛽-1a formulation to be delivered to the lungs via inhalation [619].
SNG001, which contains recombinant interferon beta-1a, was previously shown to be effective in reducing viral load in an in vivo model of swine flu and in vitro models of other coronavirus infections [620].
In July, a press release from Synairgen stated that SNG001 reduced progression to ventilation in a double-blind, placebo-controlled, multi-center study of 101 patients with an average age in the late 50s [621].
These results were subsequently published in November 2020 [622].
The study reports that the participants were assigned at a ratio of 1:1 to receive either SNG001 or a placebo that lacked the active compound, by inhalation for up to 14 days.
The primary outcome they assessed was the change in patients’ score on the WHO Ordinal Scale for Clinical Improvement (OSCI) at trial day 15 or 16.
SNG001 was associated with an odds ratio of improvement on the OSCI scale of 2.32 (95% CI 1.07 – 5.04, p = 0.033) in the intention-to-treat analysis and 2.80 (95% CI 1.21 – 6.52, p = 0.017) in the per-protocol analysis, corresponding to significant improvement in the SNG001 group on the OSCI at day 15/16.
Some of the secondary endpoints analyzed also showed differences: at day 28, the OR for clinical improvement on the OSCI was 3.15 (95% CI 1.39 – 7.14, p = 0.006), and the odds of recovery at day 15/16 and at day 28 were also significant between the two groups.
Thus, this study suggested that IFN-𝛽1 administered via SNG001 may improve clinical outcomes.
In contrast, the WHO Solidarity trial reported no significant effect of IFN-𝛽1a on patient survival during hospitalization [424].
Here, the primary outcome analyzed was in-hospital mortality, and the rate ratio for the two groups was 1.16 (95% CI, 0.96 to 1.39; p = 0.11) administering IFN-𝛽-1a to 2050 patients and comparing their response to 2,050 controls.
However, there are a few reasons that the different findings of the two trials might not speak to the underlying efficacy of this treatment strategy.
One important consideration is the stage of COVID-19 infection analyzed in each study.
The Synairgen trial enrolled only patients who were not receiving invasive ventilation, corresponding to a less severe stage of disease than many patients enrolled in the SOLIDARITY trial, as well as a lower overall rate of mortality [623].
Additionally, the methods of administration differed between the two trials, with the SOLIDARITY trial administering IFN-𝛽-1a subcutaneously [623].
The differences in findings between the studies suggests that the method of administration might be relevant to outcomes, with nebulized IFN-𝛽-1a more directly targeting receptors in the lungs.
A trial that analyzed the effect of subcutaneously administered IFN-β-1a on patients with ARDS between 2015 and 2017 had also reported no effect on 28-day mortality [624], while a smaller study analyzing the effect of subcutaneous IFN administration did find a significant improvement in 28-day mortality for COVID-19 [625].
At present, several ongoing clinical trials are investigating the potential effects of IFN-𝛽-1a, including in combination with therapeutics such as remdesivir [626] and administered via inhalation [619].
Thus, as additional information becomes available, a more detailed understanding of whether and under which circumstances IFN-𝛽-1a is beneficial to COVID-19 patients should develop.
5.6 Discussion
With the emergence of the COVID-19 pandemic caused by the coronavirus SARS-CoV-2, the development and identification of therapeutic and prophylactic interventions became issues of international urgency.
In previous outbreaks of HCoV, namely SARS and MERS, the development of these interventions was very limited.
As research has progressed, several potential approaches to treatment have emerged (Figure 4).
Most notably, remdesivir has been approved by the FDA for the treatment of COVID-19, and dexamethasone, which was approved by the FDA in 1958, has been found to improve outcomes for patients with severe COVID-19.
Other potential therapies are being still being explored and require additional data (Figure 3).
As more evidence becomes available, the potential for existing and novel therapies to improve outcomes for COVID-19 patients will become better understood.
Figure 4:Mechanism of Action for Potential Therapeutics
Potential therapeutics currently being studied can target the SARS-CoV-2 virus or modify the host environment through many different mechanisms.
Here, the relationship between the virus and several therapeutics described above are visualized.
Insights into the pathogenesis of and immune response to SARS-CoV-2 (see [1]) have also guided the identification of potential prophylactics and therapeutics.
As cases have become better characterized, it has become evident that many patients experience an initial immune response to the virus that is typically characterized by fever, cough, dyspnea, and related symptoms.
However, the most serious concern is CRS, when the body’s immune response becomes dysregulated, resulting in an extreme inflammatory response.
The RECOVERY trial, a large-scale, multi-arm trial enrolling about 15% of all COVID-19 patients in the United Kingdom, was the first to identify that the widely available steroid dexamethasone seems to be beneficial for patients suffering from this immune dysregulation [546].
The results of efforts to identify therapeutic treatments to treat patients early in the course of infection have been more ambiguous.
Early interest in the drugs HCQ and CQ yielded no promising results from studies with robust experimental designs.
On the other hand, the experimental drug remdesivir, which was developed as a candidate therapeutic for EVD, has received enough support from early analyses to receive FDA approval, although results have been mixed.
The potential for other drugs, such as tocilizumab, to reduce recovery time remains unclear, but some early results were promising.
One additional concern is that the presentation of COVID-19 appears to be heterogeneous across the lifespan.
Many adult cases, especially in younger adults, present with mild symptoms or even asymptomatically, while others, especially in older adults, can be severe or fatal.
In children, the SARS-CoV-2 viral infection can present either as a respiratory illness comparable to COVID-19 or as an inflammatory condition, known as multisystem inflammatory syndrome in children, for which presentation is similar to Kawasaki Disease [627].
The therapeutics and prophylactics discussed here were primarily tested in adults, and additional research is needed to identify therapeutics that address the symptoms characteristic of pediatric COVID-19 and MIS-C cases.
5.6.1 Potential Avenues of Interest for Therapeutic Development
Given what is currently known about these therapeutics for COVID-19, a number of related therapies beyond those explored above may also prove to be of interest.
For example, the demonstrated benefit of dexamethasone and the ongoing potential of tocilizumab for treatment of COVID-19 suggests that other anti-inflammatory agents might also hold value for the treatment of COVID-19.
Current evidence supporting the treatment of severe COVID-19 with dexamethasone suggests that the need to curtail the cytokine storm inflammatory response transcends the risks of immunosuppression, and other anti-inflammatory agents may therefore benefit patients in this phase of the disease.
While dexamethasone is considered widely available and generally affordable, the high costs of biologics such as tocilizumab therapy may present obstacles to wide-scale distribution of this drug if it proves of value.
At the doses used for RA patients, the cost for tocilizumab ranges from $179.20 to $896 per dose for the IV form and $355 for the pre-filled syringe [628].
Several other anti-inflammatory agents used for the treatment of autoimmune diseases may also be able to counter the effects of the cytokine storm induced by the virus, and some of these, such as cyclosporine, are likely to be more cost-effective and readily available than biologics [629].
While tocilizumab targets IL-6, several other inflammatory markers could be potential targets, including TNF-α.
Inhibition of TNF-α by a compound such as Etanercept was previously suggested for treatment of SARS-CoV-1 [630] and may be relevant for SARS-CoV-2 as well.
Another anti-IL-6 antibody, sarilumab, is also being investigated [631,632].
Baricitinib and other small molecule inhibitors of the Janus-activated kinase pathway also curtail the inflammatory response and have been suggested as potential options for SARS-CoV-2 infections [633].
Baricitinib, in particular, may be able to reduce the ability of SARS-CoV-2 to infect lung cells [634].
Clinical trials studying baricitinib in COVID-19 have already begun in the US and in Italy [635,636].
Identification and targeting of further inflammatory markers that are relevant in SARS-CoV-2 infection may be of value for curtailing the inflammatory response and lung damage.
In addition to immunosuppressive treatments, which are most beneficial late in disease progression, much research is focused on identifying therapeutics for early-stage patients.
For example, although studies of HCQ have not supported the early theory-driven interest in this antiviral treatment, alternative compounds with related mechanisms may still have potential.
Hydroxyferroquine derivatives of HCQ have been described as a class of bioorganometallic compounds that exert antiviral effects with some selectivity for SARS-CoV-1 in vitro[637].
Future work could explore whether such compounds exert antiviral effects against SARS-CoV-2 and whether they would be safer for use in COVID-19.
Another potential approach is the development of antivirals, which could be broad-spectrum, specific to coronaviruses, or targeted to SARS-CoV-2.
Development of new antivirals is complicated by the fact that none have yet been approved for human coronaviruses.
Intriguing new options are emerging, however.
Beta-D-N4-hydroxycytidine is an orally bioavailable ribonucleotide analog showing broad-spectrum activity against RNA viruses, which may inhibit SARS-CoV-2 replication in vitro and in vivo in mouse models of HCoVs [638].
A range of other antivirals are also in development.
Development of antivirals will be further facilitated as research reveals more information about the interaction of SARS-CoV-2 with the host cell and host cell genome, mechanisms of viral replication, mechanisms of viral assembly, and mechanisms of viral release to other cells; this can allow researchers to target specific stages and structures of the viral life cycle.
Finally, antibodies against viruses, also known as antiviral monoclonal antibodies, could be an alternative as well and are described in detail in an above section.
The goal of antiviral antibodies is to neutralize viruses through either cell-killing activity or blocking of viral replication [639].
They may also engage the host immune response, encouraging the immune system to hone in on the virus.
Given the cytokine storm that results from immune system activation in response to the virus, which has been implicated in worsening of the disease, an nAb may be preferable.
Upcoming work may explore the specificity of nAbs for their target, mechanisms by which the nAbs impede the virus, and improvements to antibody structure that may enhance the ability of the antibody to block viral activity.
Some research is also investigating potential therapeutics and prophylactics that would interact with components of the innate immune response.
For example, TLRs are pattern recognition receptors that recognize pathogen- and damage-associated molecular patterns and contribute to innate immune recognition and, more generally, promotion of both the innate and adaptive immune responses [144].
In mouse models, poly(I:C) and CpG, which are agonists of Toll-like receptors TLR3 and TLR9, respectively, showed protective effects when administered prior to SARS-CoV-1 infection [640].
Therefore, TLR agonists hold some potential for broad-spectrum prophylaxis.
Given that a large number of clinical trials are currently in progress, more information about the potential of these and other therapeutics should become available over time.
This information, combined with advances in understanding the molecular structure and viral pathogenesis of SARS-CoV-2, may lead to a more complete understanding of how the virus affects the human host and what strategies can improve outcomes.
To date, investigations of potential therapeutics for COVID-19 have focused primarily on repurposing existing drugs.
This approach is necessary given the urgency of the situation as well as the extensive time required for developing and testing new therapies.
However, in the long-term, new drugs specific for treatment of COVID-19 may also enter development.
Development of novel drugs is likely to be guided by what is known about the pathogenesis and molecular structure of SARS-CoV-2.
For example, understanding the various structural components of SARS-CoV-2 may allow for the development of small molecule inhibitors of those components.
Currently, crystal structures of the SARS-CoV-2 main protease have recently been resolved [459,641], and efforts are already in place to perform screens for small molecule inhibitors of the main protease, which have yielded potential hits [459].
Much work remains to be done to determine further crystal structures of other viral components, understand the relative utility of targeting different viral components, perform additional small molecule inhibitor screens, and determine the safety and efficacy of the potential inhibitors.
While still nascent, work in this area is promising.
Over the longer term, this approach and others may lead to the development of novel therapeutics specifically for COVID-19 and SARS-CoV-2.
5.6.2 Conclusions
Table 1: Summary table of candidate therapeutics examined in this manuscript. The FDA status is provided where available. The evidence available is based on the progression of the therapeutic through the pharmaceutical development pipeline, with randomized control trials (RCT) as the most informative source of evidence. The effectiveness is summarized based on the current available evidence; large trials such as RECOVERY and Solidarity are weighted heavily in this summary. This table was last updated on February 17, 2021.
Treatment
Category
FDA Status
Evidence Available
Suggested Effectiveness
Favipiravir
Small molecule, antiviral, nucleoside analog
None
RCT
Not supported: RCTs do not show significant improvements for individuals taking this treatment, good safety profile
Remdesivir
Small molecule, antiviral, adenosine analog
Approved for COVID-19 (and EUA for combination with baricitinib)
RCT
Conflicting evidence from large WHO-led Solidarity trial vs US-focused RCT and other studies
N3
Small molecule, protease inhibitor
None
Computational prediction, in vitro studies
Unknown
ARBs & ACEIs
Small molecule, broad spectrum
None
Observational studies and some RCTs
Not supported: Observational study retracted, RCTs suggest no association
HCQ/CQ
Small molecule, broad spectrum
None
RCT
Not supported, possibly harmful: Non-blinded RCTs showed no improvement over SOC, safety profile may be problematic
Dexamethasone
Small molecule, broad spectrum
Used off-label
RCT
Supported: RCT shows improved outcomes over SOC, especially in severe cases such as CRS
Tocilizumab
Biologic, monoclonal antibody
Approved for CRS resulting from CAR-T therapy
RCT
Mixed results from RCTs: It appears that TCZ may work well in combination with dexamethasone in severe cases, but not as monotherapy
Casirivimab and imdevimab
Biologic, monoclonal antibodies
EUA
RCT
Supported: Reduced viral load at interim analysis
Bamlanivimab and etesevimab
Biologic, monoclonal antibodies
EUA
RCT
Supported: Phase 2 clinical trial showed reduction in viral load
SNG001
Biologic, interferon
None
RCT
Mixed results: support from initial RCT but no effect found in WHO’s Solidarity trial
Due to the large number of clinical trials currently under examination (Figure 3), not all candidates are examined here (Table 1).
Instead, this review seeks to provide an overview of the range of mechanisms that have been explored and to examine some prominent candidates in the context of the pathogenesis of and immune response to SARS-CoV-2.
As more research becomes available, this review will be updated to include additional therapeutics that emerge and to include new findings that are released about those discussed here.
While no therapeutics or vaccines were developed for SARS-CoV-1 or MERS-CoV, the current state of COVID-19 research suggests that the body of literature produced before and after the emergence of these viruses has prepared the biomedical community for a rapid response to novel HCoV like SARS-CoV-2.
As the COVID-19 pandemic continues to be a topic of significant worldwide concern, more information is expected to become available about pharmaceutical mechanisms that can be used to combat this, and possibly other, HCoV.
These advances therefore not only benefit the international community’s ability to respond to the current crisis, but are also likely to shape responses to future viral threats.
6 Dietary Supplements and Nutraceuticals Under Investigation for COVID-19 Prevention and Treatment
6.1 Abstract
Coronavirus disease 2019 (COVID-19) has caused global disruption and a significant loss of life.
Existing treatments that can be repurposed as prophylactic and therapeutic agents could reduce the pandemic’s devastation.
Emerging evidence of potential applications in other therapeutic contexts has led to the investigation of dietary supplements and nutraceuticals for COVID-19.
Such products include vitamin C, vitamin D, omega 3 polyunsaturated fatty acids, probiotics, and zinc, all of which are currently under clinical investigation.
In this review, we critically appraise the evidence surrounding dietary supplements and nutraceuticals for the prophylaxis and treatment of COVID-19.
Overall, further study is required before evidence-based recommendations can be formulated, but nutritional status plays a significant role in patient outcomes, and these products could help alleviate deficiencies.
For example, evidence indicates that vitamin D deficiency may be associated with greater incidence of infection and severity of COVID-19, suggesting that vitamin D supplementation may hold prophylactic or therapeutic value.
A growing number of scientific organizations are now considering recommending vitamin D supplementation to those at high risk of COVID-19.
Because research in vitamin D and other nutraceuticals and supplements is preliminary, here we evaluate the extent to which these nutraceutical and dietary supplements hold potential in the COVID-19 crisis.
6.2 Importance
Sales of dietary supplements and nutraceuticals have increased during the pandemic due to their perceived “immune-boosting” effects.
However, little is known about the efficacy of these dietary supplements and nutraceuticals against the novel coronavirus (SARS-CoV-2) or the disease it causes, COVID-19.
This review provides a critical overview of the potential prophylactic and therapeutic value of various dietary supplements and nutraceuticals from the evidence available to date.
These include vitamin C, vitamin D, and zinc, which are often perceived by the public as treating respiratory infections or supporting immune health.
Consumers need to be aware of misinformation and false promises surrounding some supplements, which may be subject to limited regulation by authorities.
However, considerably more research is required to determine whether dietary supplements and nutraceuticals exhibit prophylactic and therapeutic value against SARS-CoV-2 infection and COVID-19.
This review provides perspective on which nutraceuticals and supplements are involved in biological processes that are relevant to recovery from or prevention of COVID-19.
6.3 Introduction
The year 2020 saw scientists and the medical community scrambling to repurpose or discover novel host-directed therapies against the coronavirus disease 2019 (COVID-19) pandemic caused by the spread of the novel Severe acute respiratory syndrome-related coronavirus 2 (SARS-CoV-2).
This rapid effort led to the identification of some promising pharmaceutical therapies for hospitalized patients, such as remdesivir and dexamethasone.
Furthermore, most societies have adopted non-pharmacological preventative measures such as utilizing public health strategies that reduce the transmission of SARS-CoV-2.
However, during this time, many individuals sought additional protections via the consumption of various dietary supplements and nutraceuticals that they believed to confer beneficial effects.
While a patient’s nutritional status does seem to play a role in COVID-19 susceptibility and outcomes [642,643,644,645,646], the beginning of the pandemic saw sales of vitamins and other supplements soar despite a lack of any evidence supporting their use against COVID-19.
In the United States, for example, dietary supplement and nutraceutical sales have shown modest annual growth in recent years (approximately 5%, or a $345 million increase in 2019), but during the six-week period preceding April 5, 2020, they increased by 44% ($435 million) relative to the same period in 2019 [647].
While growth subsequently leveled off, sales continued to boom, with a further 16% ($151 million) increase during the six weeks preceding May 17, 2020 relative to 2019 [647].
In France, New Zealand, India, and China, similar trends in sales were reported [648,649,650,651].
The increase in sales was driven by a consumer perception that dietary supplements and nutraceuticals would protect consumers from infection and/or mitigate the impact of infection due to the various “immune-boosting” claims of these products [652,653].
Due to the significant interest from the general public in dietary additives, whether and to what extent nutraceuticals or dietary supplements can provide any prophylactic or therapeutic benefit remains a topic of interest for the scientific community.
Nutraceuticals and dietary supplements are related but distinct non-pharmaceutical products.
Nutraceuticals are classified as supplements with health benefits beyond their basic nutritional value [654,655].
The key difference between a dietary supplement and a nutraceutical is that nutraceuticals should not only supplement the diet, but also aid in the prophylaxis and/or treatment of a disorder or disease [656].
However, dietary supplements and nutraceuticals, unlike pharmaceuticals, are not subject to the same regulatory protocols that protect consumers of medicines.
Indeed, nutraceuticals do not entirely fall under the responsibility of the Food and Drug Administration (FDA), but they are monitored as dietary supplements according to the Dietary Supplement, Health and Education Act 1994 (DSHEA) [657] and the Food and Drug Administration Modernization Act 1997 (FDAMA) [658].
Due to increases in sales of dietary supplements and nutraceuticals, in 1996 the FDA established the Office of Dietary Supplement Programs (ODSP) to increase surveillance.
Novel products or nutraceuticals must now submit a new dietary ingredient notification to the ODSP for review.
There are significant concerns that these legislations do not adequately protect the consumer as they ascribe responsibility to the manufacturers to ensure the safety of the product before manufacturing or marketing [659].
Manufacturers are not required to register or even seek approval from the FDA to produce or sell food supplements or nutraceuticals.
Health or nutrient content claims for labeling purposes are approved based on an authoritative statement from the Academy of Sciences or relevant federal authorities once the FDA has been notified and on the basis that the information is known to be true and not deceptive [659].
Therefore, there is often a gap between perceptions by the American public about a nutraceutical or dietary supplement and the actual clinical evidence surrounding its effects.
Despite differences in regulations, similar challenges exist outside of the United States.
In Europe, where the safety of supplements is monitored by the European Union (EU) under Directive 2002/46/EC [660].
However, nutraceuticals are not directly mentioned.
Consequently, nutraceuticals can be generally described as either a medicinal product under Directive 2004/27/EC [661] or as a ‘foodstuff’ under Directive 2002/46/EC of the European council.
In order to synchronize the various existing legislations, Regulation EC 1924/2006 on nutrition and health claims was put into effect to assure customers of safety and efficacy of products and to deliver understandable information to consumers.
However, specific legislation for nutraceuticals is still elusive.
Health claims are permitted on a product label only following compliance and authorization according to the European Food Safety Authority (EFSA) guidelines on nutrition and health claims [662].
EFSA does not currently distinguish between food supplements and nutraceuticals for health claim applications of new products, as claim authorization is dependent on the availability of clinical data in order to substantiate efficacy [663].
These guidelines seem to provide more protection to consumers than the FDA regulations but potentially at the cost of innovation in the sector [664].
The situation becomes even more complicated when comparing regulations at a global level, as countries such as China and India have existing regulatory frameworks for traditional medicines and phytomedicines not commonly consumed in Western society [665].
Currently, there is debate among scientists and regulatory authorities surrounding the development of a widespread regulatory framework to deal with the challenges of safety and health claim substantiation for nutraceuticals [659,663], as these products do not necessarily follow the same rigorous clinical trial frameworks used to approve the use of pharmaceuticals.
Such regulatory disparities have been highlighted by the pandemic, as many individuals and companies have attempted to profit from the vulnerabilities of others by overstating claims in relation to the treatment of COVID-19 using supplements and nutraceuticals.
The FDA has written several letters to prevent companies marketing or selling products based on false hyperbolic promises about preventing SARS-CoV-2 infection or treating COVID-19 [666,667,668].
These letters came in response to efforts to market nutraceutical prophylactics against COVID-19, some of which charged the consumer as much as $23,000 [669].
There have even been some incidents highlighted in the media because of their potentially life threatening consequences; for example, the use of oleandrin was touted as a potential “cure” by individuals close to the former President of the United States despite its high toxicity [670].
Thus, heterogeneous and at times relaxed regulatory standards have permitted high-profile cases of the sale of nutraceuticals and dietary supplements that are purported to provide protection against COVID-19, despite a lack of research into these compounds.
Notwithstanding the issues of poor safety, efficacy, and regulatory oversight, some dietary supplements and nutraceuticals have exhibited therapeutic and prophylactic potential.
Some have been linked with reduced immunopathology, antiviral and anti-inflammatory activities, or even the prevention of acute respiratory distress syndrome (ARDS) [652,671,672].
A host of potential candidates have been highlighted in the literature that target various aspects of the COVID-19 viral pathology, while others are thought to prime the host immune system.
These candidates include vitamins and minerals along with extracts and omega-3 polyunsaturated fatty acids (n-3 PUFA) [673].
In vitro and in vivo studies suggest that nutraceuticals containing phycocyanobilin, N-acetylcysteine, glucosamine, selenium or phase 2 inductive nutraceuticals (e.g. ferulic acid, lipoic acid, or sulforaphane) can prevent or modulate RNA virus infections via amplification of the signaling activity of mitochondrial antiviral-signaling protein (MAVS) and activation of Toll-like receptor 7 [674].
Phase 2 inductive molecules used in the production of nutraceuticals are known to activate nuclear factor erythroid 2–related factor 2 (Nrf2), which is a protein regulator of antioxidant enzymes that leads to the induction of several antioxidant enzymes, such as gamma-glutamylcysteine synthetase.
While promising, further animal and human studies are required to assess the therapeutic potential of these various nutrients and nutraceuticals against COVID-19.
For the purpose of this review, we have highlighted some of the main dietary supplements and nutraceuticals that are currently under investigation for their potential prophylactic and therapeutic applications.
These include n-3 PUFA, zinc, vitamins C and D, and probiotics.
6.4 n-3 PUFA
One category of supplements that has been explored for beneficial effects against various viral infections are the n-3 PUFAs [673], commonly referred to as omega-3 fatty acids, which include eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA).
EPA and DHA intake can come from a diet high in fish or through dietary supplementation with fish oils or purified oils [675].
Other, more sustainable sources of EPA and DHA include algae [676,677], which can also be exploited for their rich abundance of other bioactive compounds such as angiotensin converting enzyme inhibitor peptides and antiviral agents including phycobiliproteins, sulfated polysaccharides, and calcium-spirulan [678].
n-3 PUFAs have been investigated for many years for their therapeutic potential [679].
Supplementation with fish oils is generally well tolerated [679], and intake of n-3 PUFAs through dietary sources or supplementation is specifically encouraged for vulnerable groups such as pregnant and lactating women [680,681].
As a result, these well-established compounds have drawn significant interest for their potential immune effects and therapeutic potential.
Particular interest has arisen in n-3 PUFAs as potential therapeutics against diseases associated with inflammation.
n-3 PUFAs have been found to modulate inflammation by influencing processes such as leukocyte chemotaxis, adhesion molecule expression, and the production of eicosanoids [682,683].
This and other evidence indicates that n-3 PUFAs may have the capacity to modulate the adaptive immune response [655,675,682]; for example, they have been found to influence antigen presentation and the production of CD4(+) Th1 cells, among other relevant effects [684].
Certainly, preliminary evidence from banked blood samples from 100 COVID-19 patients suggests that patients with a higher omega-3 index, a measure of the amount of EPA and DHA in red blood cells, had a lower risk of death due to COVID-19 [685].
Interest has also arisen as to whether nutritional status related to n-3 PUFAs can also affect inflammation associated with severe disease, such as ARDS or sepsis [686,687].
ARDS and sepsis hold particular concern in the treatment of severe COVID-19; an analysis of 82 deceased COVID-19 patients in Wuhan during January to February 2020 reported that respiratory failure (associated with ARDS) was the cause of death in 69.5% of cases, and sepsis or multi-organ failure accounted for 28.0% of deaths [688].
Research in ARDS prior to current pandemic suggests that n-3 PUFAs may hold some therapeutic potential.
One study randomized 16 consecutive ARDS patients to receive either a fish oil-enriched lipid emulsion or a control lipid emulsion (comprised of 100% long-chain triglycerides) under a double-blinded design [689].
They reported a statistically significant reduction in leukotriene B4 levels in the group receiving the fish oil-enriched emulsion, suggesting that the fish oil supplementation may have reduced inflammation.
However, they also reported that most of their tests were not statistically significant, and therefore it seems that additional research using larger sample sizes is required.
A recent meta-analysis of 10 randomized controlled trials (RCTs) examining the effects of n-3 PUFAs on ARDS patients did not find evidence of any effect on mortality, although the effect on secondary outcomes could not be determined due to a low quality of evidence [690].
However, another meta-analysis that examined 24 RCTs studying the effects of n-3 fatty acids on sepsis, including ARDS-induced sepsis, did find support for an effect on mortality when n-3 fatty acids were administered via enteral nutrition, although a paucity of high-quality evidence again limited conclusions [691].
Therefore, despite theoretical support for an immunomodulatory effect of n-3 PUFAs in COVID-19, evidence from existing RCTs is insufficient to determine whether supplementation offers an advantage in a clinical setting that would be relevant to COVID-19.
Another potential mechanism that has led to interest in n-3 PUFAs as protective against viral infections including COVID-19 is its potential as a precursor molecule for the biosynthesis of endogenous specialized proresolving mediators (SPM), such as protectins and resolvins, that actively resolve inflammation and infection [692].
SPM have exhibited beneficial effects against a variety of lung infections, including some caused by RNA viruses [693,694].
Several mechanisms for SPM have been proposed, including preventing the release of pro-inflammatory cytokines and chemokines or increasing phagocytosis of cellular debris by macrophages [695].
In influenza, SPM promote antiviral B lymphocytic activities [696], and protectin D1 has been shown to increase survival from H1N1 viral infection in mice by affecting the viral replication machinery [697].
It has thus been hypothesized that SPM could aid in the resolution of the cytokine storm and pulmonary inflammation associated with COVID-19 [698,699].
Another theory is that some comorbidities, such as obesity, could lead to deficiencies of SPM, which could in turn be related to the occurrence of adverse outcomes for COVID-19 [700].
However, not all studies are in agreement that n-3 PUFAs or their resulting SPM are effective against infections [701].
At a minimum, the effectiveness of n-3 PUFAs against infections would be dependent on the dosage, timing, and the specific pathogens responsible [702].
On another level, there is still the question of whether fish oils can raise the levels of SPM levels upon ingestion and in response to acute inflammation in humans [703].
Currently, Karolinska University Hospital is running a trial that will measure the levels of SPM as a secondary outcome following intravenous supplementation of n-3 PUFAs in hospitalized COVID-19 patients to determine whether n-3 PUFAs provides therapeutic value [704,705].
Therefore, while this mechanism provides theoretical support for a role for n-3 PUFAs against COVID-19, experimental support is still needed.
A third possible mechanism by which n-3 PUFAs could benefit COVID-19 patients arises from the fact that some COVID-19 patients, particularly those with comorbidities, are at a significant risk of thrombotic complications including arterial and venous thrombosis [124,706].
Therefore, the use of prophylactic and therapeutic anticoagulants and antithrombotic agents is under consideration [707,708].
Considering that there is significant evidence that n-3 fatty acids and other fish oil-derived lipids possess antithrombotic properties and anti-inflammatory properties [675,709,710], they may have therapeutic value against the prothrombotic complications of COVID-19.
In particular, concerns have been raised within the medical community about using investigational therapeutics on COVID-19 patients who are already on antiplatelet therapies due to pre-existing comorbidities because the introduction of such therapeutics could lead to issues with dosing and drug choice and/or negative drug-drug interactions [707].
In such cases, dietary sources of n-3 fatty acids or other nutraceuticals with antiplatelet activities could hold particular value for reducing the risk of thrombotic complications in patients already receiving pharmaceutical antiplatelet therapies.
A new clinical trial [711] is currently recruiting COVID-19 positive patients to investigate the anti-inflammatory activity of a recently developed, highly purified nutraceutical derivative of EPA known as icosapent ethyl (VascepaTM) [712].
Other randomized controlled trials that are in the preparatory stages intend to investigate the administration of EPA and other bioactive compounds to COVID-19 positive patients in order to observe whether anti-inflammatory effects or disease state improvements occur [713,714].
Finally, while there have been studies investigating the therapeutic value of n-3 fatty acids against ARDS in humans, there is still limited evidence of their effectiveness [715].
It should be noted that the overall lack of human studies in this area means there is limited evidence as to whether these supplements could affect COVID-19 infection.
Consequently, the clinical trials that are underway and those that have been proposed will provide valuable insight into whether the anti-inflammatory potential of n-3 PUFAs and their derivatives can be beneficial to the treatment of COVID-19.
All the same, while the evidence is not present to draw conclusions about whether n-3 PUFAs will be useful in treating COVID-19, there is likely little harm associated with a diet rich in fish oils, and interest in n-3 PUFA supplementation by the general public is unlikely to have negative effects.
6.5 Zinc
Zinc is nutrient supplement that may exhibit some benefits against RNA viral infections.
Zinc is a trace metal obtained from dietary sources or supplementation and is important for the maintenance of immune cells involved in adaptive and innate immunity [716].
Supplements can be administered orally as a tablet or as a lozenge and are available in many forms, such as zinc picolinate, zinc acetate, and zinc citrate.
Zinc is also available from dietary sources including meat, seafood, nuts, seeds, legumes, and dairy.
The role of zinc in immune function has been extensively reviewed [716].
Zinc is an important signaling molecule, and zinc levels can alter host defense systems.
In inflammatory situations such as an infection, zinc can regulate leukocyte immune responses and modulate the nuclear factor kappa-light-chain-enhancer of activated B cells, thus altering cytokine production [717,718].
In particular, zinc supplementation can increase natural killer cell levels, which are important cells for host defense against viral infections [716,719].
As a result of these immune-related functions, zinc is also under consideration for possible benefits against COVID-19.
Adequate zinc intake has been associated with reduced incidence of infection [720] and antiviral immunity [721].
A randomized, double-blind, placebo-controlled trial that administered zinc supplementation to elderly subjects over the course of a year found that zinc supplementation decreased susceptibility to infection and that zinc deficiency was associated with increased susceptibility to infection [720].
Clinical trial data supports the utility of zinc to diminish the duration and severity of symptoms associated with common colds when it is provided within 24 hours of the onset of symptoms [722,723].
An observational study showed that COVID-19 patients had significantly lower zinc levels in comparison to healthy controls and that zinc-deficient COVID-19 patients (those with levels less than 80 μg/dl) tended to have more complications (70.4% vs 30.0%, p = 0.009) and potentially prolonged hospital stays (7.9 vs 5.7 days, p = 0.048) relative to patients who were not zinc deficient [724].
In coronaviruses specifically, in vitro evidence has demonstrated that the combination of zinc (Zn2+) and zinc ionophores (pyrithione) can interrupt the replication mechanisms of SARS-CoV-GFP (a fluorescently tagged SARS-CoV-1) and a variety of other RNA viruses [725,726].
Currently, there are over twenty clinical trials registered with the intention to use zinc in a preventative or therapeutic manner for COVID-19.
However, many of these trials proposed the use of zinc in conjunction with hydroxychloroquine and azithromycin [727,728,729,730], and it is not known how the lack of evidence supporting the use of hydroxychloroquine will affect investigation of zinc.
One retrospective observational study of New York University Langone hospitals in New York compared outcomes among hospitalized COVID-19 patients administered hydroxychloroquine and azithromycin with zinc sulfate (n = 411) versus hydroxychloroquine and azithromycin alone (n = 521).
Notably, zinc is the only treatment that was used in this trial that is still under consideration as a therapeutic agent due to the lack of efficacy and potential adverse events associated with hydroxychloroquine and azithromycin against COVID-19 [731,732,733].
While the addition of zinc sulfate did not affect the duration of hospitalization, the length of ICU stays or patient ventilation duration, univariate analyses indicated that zinc did increase the frequency of patients discharged and decreased the requirement for ventilation, referrals to the ICU, and mortality [734].
However, a smaller retrospective study at Hoboken University Medical Center New Jersey failed to find an association between zinc supplementation and survival of hospitalized patients [735].
Therefore, whether zinc contributes to COVID-19 recovery remains unclear.
Other trials are now investigating zinc in conjunction with other supplements such as vitamin C or n-3 PUFA [714,736].
Though there is, overall, encouraging data for zinc supplementation against the common cold and viral infections, there is currently limited evidence to suggest zinc supplementation has any beneficial effects against the current novel COVID-19; thus, the clinical trials that are currently underway will provide vital information on the efficacious use of zinc in COVID-19 prevention and/or treatment.
However, given the limited risk and the potential association between zinc deficiency and illness, maintaining a healthy diet to ensure an adequate zinc status may be advisable for individuals seeking to reduce their likelihood of infection.
6.6 Vitamin C
Vitamins B, C, D, and E have also been suggested as potential nutrient supplement interventions for COVID-19 [673,737].
In particular vitamin C has been proposed as a potential therapeutic agent against COVID-19 due to its long history of use against the common cold and other respiratory infections [738,739].
Vitamin C can be obtained via dietary sources such as fruits and vegetables or via supplementation.
Vitamin C plays a significant role in promoting immune function due to its effects on various immune cells.
It affects inflammation by modulating cytokine production, decreasing histamine levels, enhancing the differentiation and proliferation of T- and B-lymphocytes, increasing antibody levels, and protecting against the negative effects of reactive oxygen species, among other effects related to COVID-19 pathology [740,741,742].
Vitamin C is utilized by the body during viral infections, as evinced by lower concentrations in leukocytes and lower concentrations of urinary vitamin C.
Post-infection, these levels return to baseline ranges [743,744,745,746,747].
It has been shown that as little as 0.1 g/d of vitamin C can maintain normal plasma levels of vitamin C in healthy individuals, but higher doses of at least 1-3 g/d are required for critically ill patients in ICUs [748].
Indeed, vitamin C deficiency appears to be common among COVID-19 patients [749,750].
COVID-19 is also associated with the formation of microthrombi and coagulopathy [126] that contribute to its characteristic lung pathology [751], but these symptoms can be ameliorated by early infusions of vitamin C to inhibit endothelial surface P-selectin expression and platelet-endothelial adhesion [752].
Intravenous vitamin C also reduced D-dimer levels in a case study of 17 COVID-19 patients [753].
D-dimer levels are an important indicator of thrombus formation and breakdown and are notably elevated in COVID-19 patients [122,123].
There is therefore preliminary evidence suggesting that vitamin C status and vitamin C administration may be relevant to COVID-19 outcomes.
Larger-scale studies of vitamin C, however, have provided mixed results.
A recent meta-analysis found consistent support for regular vitamin C supplementation reducing the duration of the common cold, but that supplementation with vitamin C (> 200 mg) failed to reduce the incidence of colds [754].
Individual studies have found Vitamin C to reduce the susceptibility of patients to lower respiratory tract infections, such as pneumonia [755].
Another meta-analysis demonstrated that in twelve trials, vitamin C supplementation reduced the length of stay of patients in intensive care units (ICUs) by 7.8% (95% CI: 4.2% to 11.2%; p = 0.00003).
Furthermore, high doses (1-3 g/day) significantly reduced the length of an ICU stay by 8.6% in six trials (p = 0.003).
Vitamin C also shortened the duration of mechanical ventilation by 18.2% in three trials in which patients required intervention for over 24 hours (95% CI 7.7% to 27%; p = 0.001) [748].
Despite these findings, an RCT of 167 patients known as CITRUS ALI failed to show a benefit of a 96-hour infusion of vitamin C to treat ARDS [756].
Clinical trials specifically investigating vitamin C in the context of COVID-19 have now begun, as highlighted by Carr et al. [739].
These trials intend to investigate the use of intravenous vitamin C in hospitalized COVID-19 patients.
The first trial to report initial results took place in Wuhan, China [757].
These initial results indicated that the administration of 12 g/12 hr of intravenous vitamin C for 7 days in 56 critically ill COVID-19 patients resulted in a promising reduction of 28-day mortality (p = 0.06) in univariate survival analysis [758].
Indeed, the same study reported a significant decrease in IL-6 levels by day 7 of vitamin C infusion (p = 0.04) [759].
Additional studies that are being conducted in Canada, China, Iran, and the USA will provide additional insight into whether vitamin C supplementation affects COVID-19 outcomes on a larger scale.
Even though evidence supporting the use of vitamin C is beginning to emerge, we will not know how effective vitamin C is as a therapeutic for quite some time.
Currently (as of January 2021) over fifteen trials are registered with clinicaltrials.gov that are either recruiting, active or are currently in preparation.
When completed, these trials will provide crucial evidence on the efficacy of vitamin C as a therapeutic for COVID-19 infection.
However, the majority of supplementation studies investigate the intravenous infusion of vitamin C in severe patients.
Therefore, there is a lack of studies investigating the potential prophylactic administration of vitamin C via oral supplementation for healthy individuals or potentially asymptomatic SARS-CoV-2 positive patients.
Once again, vitamin C intake is part of a healthy diet and the vitamin likely presents minimal risk, but its potential prophylactic or therapeutic effects against COVID-19 are yet to be determined.
To maintain vitamin C status, it would be prudent for individuals to ensure that they consume the recommended dietary allowance of vitamin C to maintain a healthy immune system [642].
The recommended dietary allowance according to the FDA is 75-90 mg/d, whereas EFSA recommends 110 mg/d [760].
6.7 Vitamin D
Of all of the supplements currently under investigation, vitamin D has become a leading prophylactic and therapeutic candidate against SARS-CoV-2.
Vitamin D can modulate both the adaptive and innate immune system and is associated with various aspects of immune health and antiviral defense [761,762,763,764,765].
Vitamin D can be sourced through diet or supplementation, but it is mainly biosynthesized by the body on exposure to ultraviolet light (UVB) from sunlight.
Vitamin D deficiency is associated with an increased susceptibility to infection [766].
In particular, vitamin D deficient patients are at risk of developing acute respiratory infections [767] and ARDS [767].
1,25-dihydroxyvitamin D3 is the active form of vitamin D that is involved in adaptive and innate responses; however, due to its low concentration and a short half life of a few hours, vitamin D levels are typically measured by the longer lasting and more abundant precursor 25-hydroxyvitamin D.
The vitamin D receptor is expressed in various immune cells, and vitamin D is an immunomodulator of antigen presenting cells, dendritic cells, macrophages, monocytes, and T- and B-lymphocytes [766,768].
Due to its potential immunomodulating properties, vitamin D supplementation may be advantageous to maintain a healthy immune system.
Early in the pandemic it was postulated that an individual’s vitamin D status could significantly affect their risk of developing COVID-19 [769].
This hypothesis was derived from the fact that the current pandemic emerged in Wuhan China during winter, when 25-hydroxyvitamin D concentrations are at their lowest due to a lack of sunlight, whereas in the Southern Hemisphere, where it was nearing the end of the summer and higher 25-hydroxyvitamin D concentrations would be higher, the number of cases was low.
This led researchers to question whether there was a seasonal component to the SARS-CoV-2 pandemic and whether vitamin D levels might play a role [769,770,771,772].
Though it is assumed that COVID-19 is seasonal, multiple other factors that can affect vitamin D levels should also be considered.
These factors include an individual’s nutritional status, their age, their occupation, skin pigmentation, potential comorbidities, and the variation of exposure to sunlight due to latitude amongst others.
Indeed, it has been estimated that each degree of latitude north of 28 degrees corresponded to a 4.4% increase of COVID-19 mortality, indirectly linking a persons vitamin D levels via exposure to UVB light to COVID-19 mortality [770].
As the pandemic has evolved, additional research of varying quality has investigated some of the potential links identified early in the pandemic [769] between vitamin D and COVID-19.
Indeed, studies are beginning to investigate whether there is any prophylactic and/or therapeutic relationship between vitamin D and COVID-19.
A study in Switzerland demonstrated that 27 SARS-CoV-2 positive patients exhibited 25-hydroxyvitamin D plasma concentrations that were significantly lower (11.1 ng/ml) than those of SARS-CoV-2 negative patients (24.6 ng/ml; p = 0.004), an association that held when stratifying patients greater than 70 years old [773].
These findings seem to be supported by a Belgian observational study of 186 SARS-CoV-2 positive patients exhibiting symptoms of pneumonia, where 25-hydroxyvitamin D plasma concentrations were measured and CT scans of the lungs were obtained upon hospitalization [774].
A significant difference in 25-hydroxyvitamin D levels was observed between the SARS-CoV-2 patients and 2,717 season-matched hospitalized controls.
It is not clear from the study which diseases caused the control subjects to be admitted at the time of their 25-hydroxyvitamin D measurement, which makes it difficult to assess the observations reported.
Both female and male patients possessed lower median 25-hydroxyvitamin D concentrations than the control group as a whole (18.6 ng/ml versus 21.5 ng/ml; p = 0.0016) and a higher rate of vitamin D deficiency (58.6% versus 42.5%).
However, when comparisons were stratified by sex, evidence of sexual dimorphism became apparent, as female patients had equivalent levels of 25-hydroxyvitamin D to females in the control group, whereas male patients were deficient in 25-hydroxyvitamin D relative to male controls (67% versus 49%; p = 0.0006).
Notably, vitamin D deficiency was progressively lower in males with advancing radiological disease stages (p = 0.001).
However, these studies are supported by several others that indicate that vitamin D status may be an independent risk factor for the severity of COVID-19 [775,776,777,778] and in COVID-19 patients relative to population-based controls [779].
Indeed, serum concentrations of 25-hydroxyvitamin D above 30 ng/ml, which indicate vitamin D sufficiency, seems to be associated with a reduction in serum C-reactive protein, an inflammatory marker, along with increased lymphocyte levels, which suggests that vitamin D levels may modulate the immune response by reducing risk for cytokine storm in response to SARS-CoV-2 infection [779].
A study in India determined that COVID-19 fatality was higher in patients with severe COVID-19 and low serum 25-hydroxyvitamin D (mean level 6.2 ng/ml; 97% vitamin D deficient) levels versus asymptomatic non-severe patients with higher levels of vitamin D (mean level 27.9 ng/ml; 33% vitamin D deficient) [780].
In the same study, vitamin D deficiency was associated with higher levels of inflammatory markers including IL-6, ferritin, and tumor necrosis factor α.
Collectively, these studies add to a multitude of observational studies reporting potential associations between low levels of 25-hydroxyvitamin D and COVID-19 incidence and severity [773,778,779,781,782,783,784,785,786,787].
Despite the large number of studies establishing a link between vitamin D status and COVID-19 severity, an examination of data from the UK Biobank did not support this thesis [788,789].
These analyses examined 25-hydroxyvitamin D concentrations alongside SARS-CoV-2 positivity and COVID-19 mortality in over 340,000 UK Biobank participants.
However, these studies have caused considerable debate that will likely be settled following further studies [790,791].
Overall, while the evidence suggests that there is likely an association between low serum 25-hydroxyvitamin D and COVID-19 incidence, these studies must be interpreted with caution, as there is the potential for reverse causality, bias, and other confounding factors including that vitamin D deficiency is also associated with numerous pre-existing conditions and risk factors that can increase the risk for severe COVID-19 [642,770,792,793].
While these studies inform us of the potential importance of vitamin D sufficiency and the risk of SARS-CoV-2 infection and severe COVID-19, they fail to conclusively determine whether vitamin D supplementation can therapeutically affect the clinical course of COVID-19.
In one study, 40 vitamin D deficient asymptomatic or mildly symptomatic participants patients were either randomized to receive 60,000 IU of cholecalciferol daily for at least 7 days (n = 16) or a placebo (n = 24) with a target serum 25-hydroxyvitamin D level >50 ng/ml.
At day 7, 10 patients achieved >50 ng/ml, followed by another 2 by day 14.
By the end of the study, the treatment group had a greater proportion of vitamin D-deficient participants that tested negative for SARS-CoV-2 RNA, and they had a significantly lower fibrinogen levels, potentially indicating a beneficial effect [794].
A pilot study in Spain determined that early administration of high dose calcifediol (~21,000 IU days 1-2 and ~11,000 IU days 3-7 of hospital admission) with hydroxychloroquine and azithromycin to 50 hospitalized COVID-19 patients significantly reduced ICU admissions and may have reduced disease severity versus hydroxychloroquine and azithromycin alone [795].
Although this study received significant criticism from the National Institute for Health and Care Excellence (NICE) in the UK [796], an independent follow-up statistical analysis supported the findings of the study with respect to the results of cholecalciferol treatment [797].
Another trial of 986 patients hospitalized for COVID-19 in three UK hospitals administered cholecalciferol (≥ 280,000 IU in a time period of 7 weeks) to 151 patients and found an association with a reduced risk of COVID-19 mortality, regardless of baseline 25-hydroxyvitamin D levels [798].
However, a double-blind, randomized, placebo-controlled trial of 240 hospitalized COVID-19 patients in São Paulo, Brazil administered a single 200,000 IU oral dose of vitamin D.
At the end of the study, there was a 24 ng/mL difference of 25-hydroxyvitamin D levels in the treatment group versus the placebo group (p = 0.001), and 87% of the treatment group were vitamin D sufficient versus ~11% in the placebo group.
Supplementation was well tolerated.
However, there was no reduction in the length of hospital stay or mortality, and no change to any other relevant secondary outcomes were reported [799].
These early findings are thus still inconclusive with regards to the therapeutic value of vitamin D supplementation.
However, other trials are underway, including one trial that is investigating the utility of vitamin D as an immune-modulating agent by monitoring whether administration of vitamin D precipitates an improvement of health status in non-severe symptomatic COVID-19 patients and whether vitamin D prevents patient deterioration [800].
Other trials are examining various factors including mortality, symptom recovery, severity of disease, rates of ventilation, inflammatory markers such as C-reactive protein and IL-6, blood cell counts, and the prophylactic capacity of vitamin D administration [800,801,802,803].
Concomitant administration of vitamin D with pharmaceuticals such as aspirin [804] and bioactive molecules such as resveratrol [805] is also under investigation.
The effectiveness of vitamin D supplementation against COVID-19 remains open for debate.
All the same, there is no doubt that vitamin D deficiency is a widespread issue and should be addressed not only because of its potential link to SARS-CoV-2 incidence [806], but also due to its importance for overall health.
There is a possibility that safe exposure to sunlight could improve endogenous synthesis of vitamin D, potentially strengthening the immune system.
However, sun exposure is not sufficient on its own, particularly in the winter months.
Indeed, while the possible link between vitamin D status and COVID-19 is further investigated, preemptive supplementation of vitamin D and encouraging people to maintain a healthy diet for optimum vitamin D status is likely to raise serum levels of 25-hydroxyvitamin D while being unlikely to carry major health risks.
These principles seem to be the basis of a number of guidelines issued by some countries and scientific organizations that have advised supplementation of vitamin D during the pandemic.
The Académie Nationale de Médecine in France recommends rapid testing of 25-hydroxyvitamin D for people over 60 years old to identify those most at risk of vitamin D deficiency and advises them to obtain a bolus dose of 50,000 to 100,000 IU vitamin D to limit respiratory complications.
It has also recommended that those under 60 years old should take 800 to 1,000 IU daily if they receive a SARS-CoV-2 positive test [807].
In Slovenia, doctors have been advised to provide nursing home patients with vitamin D [808].
Both Public Health England and Public Health Scotland have advised members of the Black, Asian, and minority ethnic communities to supplement for vitamin D in light of evidence that they may be at higher risk for vitamin D deficiency along with other COVID-19 risk factors, a trend that has also been observed in the United States [809,810].
However, other UK scientific bodies including the NICE recommend that individuals supplement for vitamin D as per usual UK government advice but warn that people should not supplement for vitamin D solely to prevent COVID-19.
All the same, the NICE has provided guidelines for research to investigate the supplementation of vitamin D in the context of COVID-19 [811].
Despite vitamin D deficiency being a widespread issue in the United States [812], the National Institutes of Health have stated that there is “insufficient data to recommend either for or against the use of vitamin D for the prevention or treatment of COVID-19” [813].
These are just some examples of how public health guidance has responded to the emerging evidence regarding vitamin D and COVID-19.
Outside of official recommendations, there is also evidence that individuals may be paying increased attention to their vitamin D levels, as a survey of Polish consumers showed that 56% of respondents used vitamin D during the pandemic [814].
However, some companies have used the emerging evidence surrounding vitamin D to sell products that claim to prevent and treat COVID-19, which in one incident required a federal court to intervene and issue an injunction barring the sale of vitamin-D-related products due to the lack of clinical data supporting these claims [815].
It is clear that further studies and clinical trials are required to conclusively determine the prophylactic and therapeutic potential of vitamin D supplementation against COVID-19.
Until such time that sufficient evidence emerges, individuals should follow their national guidelines surrounding vitamin D intake to achieve vitamin D sufficiency.
6.8 Probiotics
Probiotics are “live microorganisms that, when administered in adequate amounts, confer a health benefit on the host” [816].
Some studies suggest that probiotics are beneficial against common viral infections, and there is modest evidence to suggest that they can modulate the immune response [817,818].
As a result, it has been hypothesized that probiotics may have therapeutic value worthy of investigation against SARS-CoV-2 [819].
Probiotics and next-generation probiotics, which are more akin to pharmacological-grade supplements, have been associated with multiple potential beneficial effects for allergies, digestive tract disorders, and even metabolic diseases through their anti-inflammatory and immunomodulatory effects [820,821].
However, the mechanisms by which probiotics affect these various conditions would likely differ among strains, with the ultimate effect of the probiotic depending on the heterogeneous set of bacteria present [821].
Some of the beneficial effects of probiotics include reducing inflammation by promoting the expression of anti-inflammatory mediators, inhibiting Toll-like receptors 2 and 4, competing directly with pathogens, synthesizing antimicrobial substances or other metabolites, improving intestinal barrier function, and/or favorably altering the gut microbiota and the brain-gut axis [821,822,823].
It is also thought that lactobacilli such as Lactobacillus paracasei, Lactobacillus plantarum and Lactobacillus rhamnosus have the capacity to bind to and inactivate some viruses via adsorptive and/or trapping mechanisms [824].
Other probiotic lactobacilli and even non-viable bacterium-like particles have been shown to reduce both viral attachment to host cells and viral titers, along with reducing cytokine synthesis, enhancing the antiviral IFN-α response, and inducing various other antiviral mechanisms [824,825,826,827,828,829,830,831,832].
These antiviral and immunobiotic mechanisms and others have been reviewed in detail elsewhere [672,819,833].
However, there is also a bi-directional relationship between the lungs and gut microbiota known as the gut-lung axis [834], whereby gut microbial metabolites and endotoxins may affect the lungs via the circulatory system and the lung microbiota in return may affect the gut [835].
Therefore, the gut-lung axis may play role in our future understanding of COVID-19 pathogenesis and become a target for probiotic treatments [836].
Moreover, as microbial dysbiosis of the respiratory tract and gut may play a role in some viral infections, it has been suggested that SARS-CoV-2 may interact with our commensal microbiota [672; 837; 10.3389/fmicb.2020.01840] and that the lung microbiome could play a role in developing immunity to viral infections [838].
These postulations, if correct, could lead to the development of novel probiotic and prebiotic treatments.
However, significant research is required to confirm these associations and their relevance to patient care, if any.
Probiotic therapies and prophylactics may also confer some advantages for managing symptoms of COVID-19 or risks associated with its treatment.
Probiotics have tentatively been associated with the reduction of risk and duration of viral upper respiratory tract infections [839,840,841].
Some meta-analyses that have assessed the efficacy of probiotics in viral respiratory infections have reported moderate reductions in the incidence and duration of infection [840,842].
Indeed, randomized controlled trials have shown that administering Bacillus subtilis and Enterococcus faecalis[843], Lactobacillus rhamnosus GG[844], or Lactobacillus casei and Bifidobacterium breve with galactooligosaccharides [845] via the nasogastric tube to ventilated patients reduced the occurrence of ventilator-associated pneumonia in comparison to the respective control groups in studies of viral infections and sepsis.
These findings were also supported by a recent meta-analysis [846].
Additionally, COVID-19 patients carry a significant risk of ventilator-associated bacterial pneumonia [847], but it can be challenging for clinicians to diagnose this infection due to the fact that severe COVID-19 infection presents with the symptoms of pneumonia [848].
Therefore, an effective prophylactic therapy for ventilator-associated pneumonia in severe COVID-19 patients would carry significant therapeutic value.
Additionally, in recent years, probiotics have become almost synonymous with the treatment of gastrointestinal issues due to their supposed anti-inflammatory and immunomodulatory effects [849].
Notably, gastrointestinal symptoms commonly occur in COVID-19 patients [850], and angiotensin-converting enzyme 2, the portal by which SARS-CoV-2 enters human cells, is highly expressed in enterocytes of the ileum and colon, suggesting that these organs may be a potential route of infection [851,852].
Indeed, SARS-CoV-2 viral RNA has been detected in human feces [105,853], and fecal-oral transmission of the virus has not yet been ruled out [854].
Rectal swabs of some SARS-CoV-2 positive pediatric patients persistently tested positive for several days despite negative nasopharyngeal tests, indicating the potential for fecal viral shedding [855].
However, there is conflicting evidence for the therapeutic value of various probiotics against the incidence or severity of gastrointestinal symptoms in viral or bacterial infections such as gastroenteritis [856,857].
Nevertheless, it has been proposed that the administration of probiotics to COVID-19 patients and healthcare workers may prevent or ameliorate the gastrointestinal symptoms of COVID-19, a hypothesis that several clinical trials are now preparing to investigate [858,859].
Other studies are investigating whether probiotics may affect patient outcomes following SARS-CoV-2 infection [860].
Generally, the efficacy of probiotic use is a controversial topic among scientists.
In Europe, EFSA has banned the term probiotics on products labels, which has elicited either criticism for EFSA or support for probiotics from researchers in the field [816,861,862].
This regulation is due to the hyperbolic claims placed on the labels of various probiotic products, which lack rigorous scientific data to support their efficacy.
Overall, the data supporting probiotics in the treatment or prevention of many different disorders and diseases is not conclusive, as the quality of the evidence is generally considered low [839].
However, in the case of probiotics and respiratory infections, the evidence seems to be supportive of their potential therapeutic value.
Consequently, several investigations are underway to investigate the prophylactic and therapeutic potential of probiotics for COVID-19.
The blind use of conventional probiotics for COVID-19 is currently cautioned against until the pathogenesis of SARS-CoV-2 can be further established [863].
Until clinical trials investigating the prophylactic and therapeutic potential of probiotics for COVID-19 are complete, it is not possible to provide an evidence-based recommendation for their use.
Despite these concerns, complementary use of probiotics as an adjuvant therapeutic has been proposed by the Chinese National Health Commission and National Administration of Traditional Chinese Medicine [106].
While supply issues prevented the probiotics market from showing the same rapid response to the COVID-19 as some other supplements, many suppliers are reporting growth during the pandemic [864].
Therefore, the public response once again seems to have adopted supplements promoted as bolstering the immune response despite a lack of evidence suggesting they are beneficial for preventing or mitigating COVID-19.
6.9 Discussion
In this review, we report the findings to date of analyses of several dietary supplements and nutraceuticals.
While existing evidence suggests potential benefits of n-3 PUFA and probiotic supplementation for COVID-19 treatment and prophylaxis, clinical data is still lacking, although trials are underway.
Both zinc and vitamin C supplementation in hospitalized patients seem to be associated with positive outcomes; however, further clinical trials are required.
In any case, vitamin C and zinc intake are part of a healthy diet and likely present minimal risk when supplemented, though their potential prophylactic or therapeutic effects against COVID-19 are yet to be determined.
On the other hand, mounting evidence from observational studies indicates that there is an association between vitamin D deficiency and COVID-19 incidence has also been supported by meta-analysis [865].
Indeed, scientists are working to confirm these findings and to determine whether a patient’s serum 25-hydroxyvitamin D levels are also associated with COVID-19 severity.
Clinical trials are required to determine whether preemptive vitamin D supplementation may mitigate against severe COVID-19.
In terms of the therapeutic potential of vitamin D, initial evidence from clinical trials is conflicting but seems to indicate that vitamin D supplementation may reduce COVID-19 severity [795].
The various clinical trials currently underway will be imperative to provide information on the efficacious use of vitamin D supplementation for COVID-19 prevention and/or treatment.
The purported prophylactic and therapeutic benefits of dietary supplements and nutraceuticals for multiple disorders, diseases, and infections has been the subject of significant research and debate for the last few decades.
Inevitably, scientists are also investigating the potential for these various products to treat or prevent COVID-19.
This interest also extends to consumers, which led to a remarkable increase of sales of dietary supplements and nutraceuticals throughout the pandemic due to a desire to obtain additional protections from infection and disease.
The nutraceuticals discussed in this review, namely vitamin C, vitamin D, n-3 PUFA, zinc, and probiotics, were selected because of potential biological mechanisms that could beneficially affect viral and respiratory infections and because they are currently under clinical investigation.
Specifically, these compounds have all been found to influence cellular processes related to inflammation.
Inflammation is particularly relevant to COVID-19 because of the negative outcomes (often death) observed in a large number of patients whose immune response becomes hyperactive in response to SARS-CoV-2, leading to severe outcomes such as ARDS and sepsis [866].
Additionally, there is a well-established link between diet and inflammation [867], potentially mediated in part by the microbiome [868].
Thus, the idea that dietary modifications or supplementation could be used to modify the inflammatory response is tied to a broader view of how diet and the immune system are interconnected.
The supplements and nutraceuticals discussed here therefore lie in sharp contrast to other alleged nutraceutical or dietary supplements that have attracted during the pandemic, such as colloidal silver [869], which have no known nutritional function and can be harmful.
Importantly, while little clinical evidence is available about the effects of any supplements against COVID-19, the risks associated with those discussed above are likely to be low, and in some cases, they can be obtained from dietary sources alone.
There are various other products and molecules that have garnered scientific interest and could merit further investigation.
These include polyphenols, lipid extracts, and tomato-based nutraceuticals, all of which have been suggested for the potential prevention of cardiovascular complications of COVID-19 such as thrombosis [672,708].
Melatonin is another supplement that has been identified as a potential antiviral agent against SARS-CoV-2 using computational methods [870], and it has also been highlighted as a potential therapeutic agent for COVID-19 due to its documented antioxidant, anti-apoptotic, immunomodulatory, and anti-inflammatory effects [708,871,872].
Notably, melatonin, vitamin D and zinc have attracted public attention because they were included in the treatment plan of the former President of the United States upon his hospitalization due to COVID-19 [873].
These are just some of the many substances and supplements that are currently under investigation but as of yet lack evidence to support their use for the prevention or treatment of COVID-19.
While there is plenty of skepticism put forward by physicians and scientists surrounding the use of supplements, these statements have not stopped consumers from purchasing these products, with one study reporting that online searches for dietary supplements in Poland began trending with the start of the pandemic [814].
Additionally, supplement usage increased between the first and second wave of the pandemic.
Participants reported various reasons for their use of supplements, including to improve immunity (60%), to improve overall health (57%), and to fill nutrient gaps in their diet (53%).
Other efforts to collect large datasets regarding such behavior have also sought to explore a possible association between vitamin or supplement consumption and COVID-19.
An observational analysis of survey responses from 327,720 users of the COVID Symptom Study App found that the consumption of n-3 PUFA supplements, probiotics, multivitamins, and vitamin D was associated with a lower risk of SARS-CoV-2 infection in women but not men after adjusting for potential confounders [874].
According to the authors, the sexual dimorphism observed may in part be because supplements may better support females due to known differences between the male and female immune systems, or it could be due to behavioral and health consciousness differences between the sexes [874].
Certainly, randomized controlled trials are required to investigate these findings further.
Finally, it is known that a patient’s nutritional status affects health outcomes in various infectious diseases [646], and COVID-19 is no different [644,875,876].
Some of the main risk factors for severe COVID-19, which also happen to be linked to poor nutritional status, include obesity, hypertension, cardiovascular diseases, type II diabetes mellitus, and indeed age-related malnutrition [642,644,877].
Although not the main focus of this review, it is important to consider the nutritional challenges associated with severe COVID-19 patients.
Hospitalized COVID-19 patients tend to report an unusually high loss of appetite preceding admission, some suffer diarrhea and gastrointestinal symptoms that result in significantly lower food intake, and patients with poorer nutritional status were more likely to have worse outcomes and require nutrition therapy [878].
Dysphagia also seems to be a significant problem in pediatric patients that suffered multisystem inflammatory syndrome [879] and rehabilitating COVID-19 patients, potentially contributing to poor nutritional status [880].
Almost two-thirds of discharged COVID-19 ICU patients exhibit significant weight loss, of which 26% had weight loss greater than 10% [876].
As investigated in this review, hospitalized patients also tend to exhibit vitamin D deficiency or insufficiency, which may be associated with greater disease severity [865].
Therefore, further research is required to determine how dietary supplements and nutraceuticals may contribute to the treatment of severely ill and rehabilitating patients, who often rely on enteral nutrition.
6.10 Conclusions
Despite all the potential benefits of nutraceutical and dietary supplement interventions presented, currently there is a paucity of clinical evidence to support their use for the prevention or mitigation of COVID-19 infection.
Nevertheless, optimal nutritional status can prime an individual’s immune system to protect against the effects of acute respiratory viral infections by supporting normal maintenance of the immune system [642,646].
Nutritional strategies can also play a role in the treatment of hospitalized patients, as malnutrition is a risk to COVID-19 patients [880].
Overall, supplementation of vitamin C, vitamin D, and zinc may be an effective method of ensuring their adequate intake to maintain optimal immune function, which may also convey beneficial effects against viral infections due to their immunomodulatory effects.
Individuals should pay attention to their nutritional status, particularly their intake of vitamin D, considering that vitamin D deficiency is widespread.
The prevailing evidence seems to indicate an association between vitamin D deficiency with COVID-19 incidence and, potentially, severity [770].
As a result, some international authorities have advised the general public, particularly those at high risk of infection, to consider vitamin D supplementation.
However, further well-controlled clinical trials are required to confirm these observations.
Many supplements and nutraceuticals designed for various ailments that are available in the United States and beyond are not strictly regulated [881].
Consequently, there can be safety and efficacy concerns associated with many of these products.
Often, the vulnerable members of society can be exploited in this regard and, unfortunately, the COVID-19 pandemic has proven no different.
As mentioned above, the FDA has issued warnings to several companies for advertising falsified claims in relation to the preventative and therapeutic capabilities of their products against COVID-19 [882].
Further intensive investigation is required to establish the effects of these nutraceuticals, if any, against COVID-19.
Until more effective therapeutics are established, the most effective mitigation strategies consist of encouraging standard public health practices such as regular hand washing with soap, wearing a face mask, and covering a cough with your elbow [883], along with following social distancing measures, “stay at home” guidelines, expansive testing, and contact tracing [884,885].
Indeed, in light of this review, it would also be pertinent to adopt a healthy diet and lifestyle following national guidelines in order to maintain optimal immune health.
Because of the broad public appeal of dietary supplements and nutraceuticals, it is important to evaluate the evidence regarding the use of such products.
We will continue to update this review as more findings become available.
7 Vaccine Development Strategies for SARS-CoV-2
7.1 Abstract
The release of the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) genome sequence on January 10th, 2020, mobilized global vaccine development on a scale never before seen in response to the coronavirus disease 2019 (COVID-19) pandemic.
Pre-existing vaccine platforms and novel vaccine technologies were deployed by manufacturers early in the pandemic.
Regardless of the platform used, almost all the vaccine candidates have targeted the spike protein of SARS-CoV-2.
A historically slow process, vaccine development accelerated to the point that less than a year later, some vaccine candidates had reported interim phase III clinical trial data and received emergency use authorization (EUA) in the United States and approval by the European Union.
In this review, the strategies used to develop the leading vaccine candidates are discussed and their safety and efficacy are appraised.
As of March 2021, publicly available safety and efficacy data generated from the phase III clinical trials supports the distribution of spike targeting adenoviral vector vaccines and mRNA vaccines.
Lastly, the initial state of global vaccine distribution is also reviewed.
7.2 Importance
SARS-CoV-2 has infected over 125 million people and cost the lives of 2.8 million people globally.
The development, production, and distribution of prophylactic vaccines is imperative to saving lives, preventing illness, and reducing the economic and social burdens caused by the COVID-19 pandemic.
Assuming effective deployment, these vaccines offer an opportunity to move into a new phase of the pandemic where the susceptibility of worldwide populations is significantly reduced.
This review highlights the main strategies utilized for the development of the COVID-19 vaccines, their clinical appraisal, and their distribution.
7.3 Introduction
For a highly infectious virus like SARS-CoV-2, a vaccine would hold particular value because it could bolster the immune response to the virus of the population broadly, thereby driving a lower rate of infection and likely significantly reducing fatalities.
However, the vaccine development process has historically been slow, and vaccines fail to provide immediate prophylactic protection or treat ongoing infections [886].
Flu-like illnesses caused by viruses are a common target of vaccine development programs, and influenza vaccine technology in particular has made many strides.
During the H1N1 influenza outbreak, vaccine development was accelerated because of the existing infrastructure, along with the fact that regulatory agencies had already decided that vaccines produced using egg- and cell-based platforms could be licensed under the regulations used for a strain change.
Critiques of the production and distribution of the H1N1 vaccine have stressed the need for alternative development-and-manufacturing platforms that can be readily adapted to new pathogens.
Although a monovalent H1N1 vaccine was not available before the pandemic peaked in the United States and Europe, it was available soon afterward as a stand-alone vaccine that was eventually incorporated into commercially available seasonal influenza vaccines [887].
If H1N1 vaccine development provides any indication, considering developing and manufacturing platforms for promising COVID-19 vaccine trials early could hasten the emergence of an effective prophylactic vaccine against SARS-CoV-2.
Figure 5:Structure of the SARS-CoV-2 virus.
The development of vaccines depends on the immune system recognizing the virus.
Here, the structure of SARS-CoV-2 is represented both in the abstract and against a visualization of the virion.
The abstracted visualization was made using BioRender [888] and the microscopy was done by the National Institute of Allergy and Infectious Diseases [889].
The first critical step towards developing a vaccine against SARS-CoV-2 was characterizing the target, which fortunately happened early in the COVID-19 outbreak with the sequencing and dissemination of the viral genome [890].
The Coalition for Epidemic Preparedness Innovations (CEPI) is coordinating global health agencies and pharmaceutical companies to develop vaccines against SARS-CoV-2.
As of September 2020, there were over 180 vaccine candidates against SARS-CoV-2 in development [891].
Unlike many global vaccine development programs previously, such as with H1N1, the vaccine development landscape for COVID-19 includes vaccines produced by a wide array of technologies.
Experience in the field of oncology is encouraging COVID-19 vaccine developers to use next-generation approaches to vaccine development, which have led to the great diversity of vaccine development programs [892].
These diverse technology platforms include DNA, RNA, virus-like particle, recombinant protein, both replicating and non-replicating viral vectors, live attenuated virus, and inactivated virus approaches (Figure 6).
Given the wide range of vaccines under development, it is possible that some vaccine products may eventually be shown to be more effective in certain subpopulations, such as children, pregnant women, immunocompromised patients, the elderly, etc.
The requirements for a successful vaccine trial and deployment are complex and may require coordination between government, industry, academia, and philanthropic entities [893].
While little is currently known about immunity to SARS-CoV-2, vaccine development typically tests for serum neutralizing activity, as this has been established as a biomarker for adaptive immunity in other respiratory illnesses [894].
Figure 6:Vaccine Development Strategies.
Several different strategies can and are being employed for the development of vaccines.
Each approach capitalizes on different features of the SARS-CoV-2 virus and delivery through a different platform.
7.4 DNA Vaccines
This vaccination method involves the direct introduction of a plasmid containing a DNA sequence encoding the antigen(s) against which an immune response is sought into appropriate tissues [895].
This approach may offer several advantages over traditional vaccination approaches, such as the stimulation of both B- as well as T-cell responses and the absence of any infectious agent.
7.4.1 INO-4800 Vaccine
Currently, a Phase I safety and immunogenicity clinical trial of INO-4800, a prophylactic vaccine against SARS-CoV-2, is underway [896].
The vaccine developer Inovio Pharmaceuticals Technology is overseeing administration of INO-4800 by intradermal injection followed by electroporation with the CELLECTRA® device to healthy volunteers.
Electroporation is the application of brief electric pulses to tissues in order to permeabilize cell membranes in a transient and reversible manner.
It has been shown that electroporation can enhance vaccine efficacy by up to 100-fold, as measured by increases in antigen-specific antibody titers [897].
The safety of the CELLECTRA® device has been studied for over seven years, and these studies support the further development of electroporation as a safe vaccine delivery method [898].
The temporary formation of pores through electroporation facilitates the successful transportation of macromolecules into cells, allowing cells to robustly take up INO-4800 for the production of an antibody response.
Approved by the U.S. FDA on April 6, 2020, the Phase I study is enrolling up to 40 healthy adult volunteers in Philadelphia, PA at the Perelman School of Medicine and at the Center for Pharmaceutical Research in Kansas City, MO.
The trial has two experimental arms corresponding to the two locations.
Participants in Experimental Group 1 will receive one intradermal injection of 1.0 milligram (mg) of INO-4800 followed by electroporation using the CELLECTRA® 2000 device twice, administered at Day 0 and Week 4.
Participants in Experimental Group 2 will receive two intradermal injections of 1.0 mg (total 2.0 mg per dosing visit) of INO-4800 followed by electroporation using the CELLECTRA® 2000 device, administered at Day 0 and Week 4.
Safety data and the initial immune responses of participants from the trial are expected by the end of the summer of 2021.
The development of a DNA vaccine against SARS-CoV-2 by Inovio could be an important step forward in the world’s search for a COVID-19 vaccine.
Although exciting, the cost of vaccine manufacturing and electroporation may make scaling the use of this technology for prophylactic use for the general public difficult.
7.5 RNA Vaccines
RNA vaccines are nucleic-acid based modalities that code for viral antigens against which the human body elicits a humoral and cellular immune response.
The mRNA technology is transcribed in vitro and delivered to cells via lipid nanoparticles (LNP) [899].
They are recognized by ribosomes in vivo and then translated and modified into functional proteins [900].
The resulting intracellular viral proteins are displayed on surface MHC proteins, provoking a strong CD8+ T cell response as well as a CD4+ T cell and B cell-associated antibody responses [900].
Naturally, mRNA is not very stable and can degrade quickly in the extracellular environment or the cytoplasm.
The LNP covering protects the mRNA from enzymatic degradation outside of the cell [901].
Codon optimization to prevent secondary structure formation and modifications of the poly-A tail as well as the 5’ untranslated region to promote ribosomal complex binding can increase mRNA expression in cells.
Furthermore, purifying out double-stranded RNA and immature RNA with FPLC (fast performance liquid chromatography) and HPLC (high performance liquid chromatography) technology will improve translation of the mRNA in the cell [900,902].
Vaccines based on mRNA delivery confer many advantages over traditional viral vectored vaccines and DNA vaccines.
In comparison to live attenuated viruses, mRNA vaccines are non-infectious and can be synthetically produced in an egg-free, cell-free environment, thereby reducing the risk of a detrimental immune response in the host [903].
Unlike DNA vaccines, mRNA technologies are naturally degradable and non-integrating, and they do not need to cross the nuclear membrane in addition to the plasma membrane for their effects to be seen [900].
Furthermore, mRNA vaccines are easily, affordably, and rapidly scalable.
Although mRNA vaccines have been developed for therapeutic and prophylactic purposes, none have previously been licensed or commercialized.
Nevertheless, they have shown promise in animal models and preliminary clinical trials for several indications, including rabies, coronavirus, influenza, and cytomegalovirus [904].
Preclinical data previously identified effective antibody generation against full-length FPLC-purified influenza hemagglutinin stalk-encoding mRNA in mice, rabbits, and ferrets [905].
Similar immunological responses for mRNA vaccines were observed in humans in Phase I and II clinical trials operated by the pharmaceutical-development companies Curevac and Moderna for rabies, flu, and zika [902].
Positively charged bilayer LNPs carrying the mRNA attract negatively charged cell membranes, endocytose into the cytoplasm [901], and facilitate endosomal escape.
LNPs can be coated with modalities recognized and engulfed by specific cell types, and LNPs that are 150 nm or less effectively enter into lymphatic vessels [901,906].
Therefore, this technology holds great potential for targeted delivery of modified mRNA.
There are three types of RNA vaccines: non-replicating, in vivo self-replicating, and in vitro dendritic cell non-replicating [907].
Non-replicating mRNA vaccines consist of a simple open reading frame (ORF) for the viral antigen flanked by the 5’ UTR and 3’ poly-A tail.
In vivo self-replicating vaccines encode a modified viral genome derived from single-stranded, positive sense RNA alphaviruses [900,902].
The RNA genome encodes the viral antigen along with proteins of the genome replication machinery, including an RNA polymerase.
Structural proteins required for viral assembly are not included in the engineered genome [900].
Self-replicating vaccines produce more viral antigens over a longer period of time, thereby evoking a more robust immune response [907].
Finally, in vitro dendritic cell non-replicating RNA vaccines limit transfection to dendritic cells.
Dendritic cells are potent antigen-presenting immune cells that easily take up mRNA and present fragments of the translated peptide on their MHC proteins, which can then interact with T cell receptors.
Ultimately, primed T follicular helper cells can stimulate germinal center B cells that also present the viral antigen to produce antibodies against the virus [908].
These cells are isolated from the patient, grown and transfected ex vivo, and reintroduced to the patient [909].
Given the potential for this technology to be quickly adapted for a new pathogen, it has held significant interest for the treatment of COVID-19.
In the vaccines developed under this approach, the spike protein, which is immunogenic [42], can be furnished to the immune system in order to train its response.
The vaccine candidates developed against SARS-CoV-2 using mRNA vectors utilize similar principles and technologies, although there are slight differences in implementation among candidates such as the formulation of the platform and the specific components of the spike protein encapsulated (e.g., the full Spike protein vs. the RBD alone) [910].
The results of the interim analyses of two mRNA vaccine candidates became available at the end of 2020 and provided strong support for this emerging approach to vaccination.
Below we describe the results available as of February 2021 for two such candidates, mRNA-1273 produced by ModernaTX and BNT162b2 produced by Pfizer, Inc. and BioNTech.
7.5.1 ModernaTX mRNA Vaccine
ModernaTX’s mRNA-1273 vaccine, which is an was the first COVID-19 vaccine to enter a phase I clinical trial in the United States.
In this trial, Moderna spearheaded an investigation on the immunogenicity and reactogenicity of mRNA-1273, a conventional lipid nanoparticle encapsulated RNA encoding a full-length prefusion stabilized S protein for SARS-CoV-2 [911].
An initial report described the results of enrolling forty-five participants who were administered intramuscular injections of mRNA-1273 in their deltoid muscle on day 1 and day 29, and then followed for the next twelve months [894].
Healthy males and non-pregnant females aged 18-55 years were recruited for this study and divided into three groups receiving 25, 100, or 250 micrograms (μg) of mRNA-1273.
IgG ELISA assays on patient serology samples were used to examine the immunogenicity of the vaccine [911].
Binding antibodies were observed at two weeks after the first dose at all concentrations.
At the time point one week after the second dose was administered on day 29, the pseudotyped lentivirus reporter single-round-of-infection neutralization assay (PsVNA), which was used to assess neutralizing activity, reached a median level similar to the median observed in convalescent plasma samples.
Participants reported mild and moderate systemic adverse events after the day 1 injection, and one severe local event was observed in each of the two highest dose levels.
The second injection led to severe systemic adverse events for three of the participants at the highest dose levels, with one participant in the group being evaluated at an urgent care center on the day after the second dose.
The reported localized adverse events from the second dose were similar to those from the first.
Several months later, a press release from ModernaTX described the results of the first interim analysis of the vaccine [912].
On November 16, 2020, a report was released describing the initial results from Phase III testing, corresponding to the first 95 cases of COVID-19 in the 30,000 enrolled participants [912], with additional data released to the FDA on December 17, 2020 [913].
These results were subsequently published in a peer-reviewed journal (The New England Journal of Medicine) on December 30, 2020 [914].
The first group of 30,420 study participants were randomized to receive the vaccine or a placebo at a ratio of 1:1 [914].
Administration occurred at 99 sites within the United States in two sessions, spaced 28 days apart [914,915].
Patients reporting COVID-19 symptoms upon follow-up were tested for SARS-CoV-2 using a nasopharyngeal swab that was evaluated with RT-PCR [915].
The initial preliminary analysis reported the results of the cases observed up until a cut-off date of November 11, 2020.
Of these first 95 cases reported, 90 occurred in participants receiving the placebo compared to 5 cases in the group receiving the vaccine [912].
These results suggested the vaccine is 94.5% effective in preventing COVID-19.
Additionally, eleven severe cases of COVID-19 were observed, and all eleven occurred in participants receiving the placebo.
The publication reported the results through an extended cut-off date of November 21, 2020, corresponding to 196 cases [914].
Of these, 11 occurred in the vaccine group and 185 in the placebo group, corresponding to an efficacy of 94.1%.
Once again, all of the severe cases of COVID-19 observed (n=30) occurred in the placebo group, including one death.
Thus, as more cases are reported, the efficacy of the vaccine has remained above 90%, and no cases of severe COVID-19 have yet been reported in participants receiving the vaccine.
These findings suggest the possibility that the vaccine might bolster immune defenses even for subjects who do still develop a SARS-CoV-2 infection.
The study was designed with an explicit goal of including individuals at high risk for COVID-19, including older adults, people with underlying health conditions, and people of color [916].
The Phase III trial population was comprised by approximately 25.3% adults over age 65 in the initial report and 24.8% in the publication [915].
Among the cases reported by both interim analyses, 16-17% occurred in older adults [912,914]..
Additionally, approximately 10% of participants identified a Black or African-American background and 20% identified Hispanic or Latino ethnicity [914,915].
Among the first 95 cases, 12.6% occurred in participants identifying a Hispanic or Latino background and 4% in participants reporting a Black or African-American background [912]; in the publication, they indicated only that 41 of the cases reported in the placebo group and 1 case in the treatment group occurred in “communities of color”, corresponding to 21.4% of all cases [914].
While the sample size in both analyses is small relative to the study population of over 30,000, these results suggest that the vaccine is likely to be effective in people from a variety of backgrounds.
By all indications, this vaccine is likely to be highly useful in mitigating the damage of SARS-CoV-2.
In-depth safety data was released by ModernaTX as part of their application for an EUA from the FDA and summarized in the associated publication [914,915].
Because the detail provided in the report is greater than that provided in the publication, here we emphasize the results observed at the time of the first analysis.
Overall, a large percentage of participants reported adverse effects when solicited, and these reports were higher in the vaccine group than in the placebo group (94.5% versus 59.5%, respectively, at the time of the initial analysis) [915].
Some of these events met the criteria for grade 3 (local or systemic) or grade 4 (systemic only) toxicity [915], but most were grade 1 or grade 2 and lasted 2-3 days [914].
The most common local adverse reaction was pain at the injection site, reported by 83.7% of participants receiving the first dose of the vaccine and 88.4% upon receiving the second dose, compared to 19.8% and 19.8% and 17.0%, respectively, of patients in the placebo condition [915].
Fewer than 5% of vaccine recipients reported grade 3 pain at either administration.
Other frequent local reactions included erythema, swelling, and lymphadenopathy [915].
For systemic adverse reactions, fatigue was the most common [915].
Among participants receiving either dose of the vaccine, 68.5% reported fatigue compared to 36.1% participants receiving the placebo [915].
The level of fatigue experienced was usually fairly mild, with only 9.6% and 1.3% of participants in the vaccine and placebo conditions, respectively, reporting grade 3 fatigue [915], which corresponds to significant interference with daily activity [917].
Based on the results of the report, an EUA was issued on December 18, 2020 to allow distribution of this vaccine in the United States [918], and it was shortly followed by an Interim Order authorizing distribution of the vaccine in Canada [919] and a conditional marketing authorization by the European Medicines Agency to facilitate distribution in the European Union [920].
7.5.2 Pfizer/BioNTech BNT162b2
ModernaTX was, in fact, the second company to release news of a successful interim analysis of an mRNA vaccine and receive an EUA.
The first report came from Pfizer and BioNTech’s mRNA vaccine BNT162b2 on November 9, 2020 [921], and a preliminary report was published in the New England Journal of Medicine one month later [329].
The vaccine candidate is contains the full prefusion stabilized, membrane-anchored SARS-CoV-2 spike protein in a vaccine formulation based on modified mRNA (modRNA) technology [922,923].
This vaccine candidate should not be confused with a similar candidate from Pfizer/BioNTech, BNT162b1, that delivered only the RBD of the spike protein [924,925], which was not advanced to a stage III trial because of the improved reactogenicity/immunogenicity profile of BNT162b2 [330].
During the Phase III trial of BNT162b2, 43,538 participants were enrolled 1:1 in the placebo and the vaccine candidate and received two 30-μg doses 21 days apart [329].
Of these enrolled participants, 21,720 received BNT162b2 and 21,728 received a placebo [329].
Recruitment occurred at 135 sites across six countries: Argentina, Brazil, Germany, South Africa, Turkey, and the United States.
An initial press release described the first 94 cases, which were consistent with 90% efficacy of the vaccine at 7 days following the second dose [921].
The release of the full trial information covered a longer period and analyzed the first 170 cases occurring at least 7 days after the second dose, 8 of which occurred in patients who had received BNT162b2.
The press release characterized the study population as diverse, reporting that 42% of the participants worldwide came from non-white backgrounds, including 10% Black and 26% Hispanic or Latino [926].
Within the United States, 10% and 13% of participants, respectively, identified themselves as having Black or Hispanic/Latino backgrounds [926].
Additionally, 41% of participants worldwide were 56 years of age or older [926], and they reported that the efficacy of the vaccine in adults over 65 was 94% [927].
The primary efficacy analysis of the Phase III study was concluded on November 18, 2020 [927], and the final results indicted 94.6% efficacy of the vaccine [329]v.
The safety profile of the vaccine was also assessed [329].
A subset of patients were followed for reactogenicity using electronic diaries, with the data collected from these 8,183 participants comprising the solicited safety events analyzed.
Much like those who received the ModernaTX vaccine candidate, a large proportion of participants reported experiencing site injection pain within 7 days of vaccination.
While percentages are broken down by age group in the publication, these proportions correspond to approximately 78% and 73% of all participants after the first and second doses, respectively, overall.
Only a small percentage of these events (less than 1%) were rated as serious, with the rest being mild or moderate, and none reached grade 4.
Some participants also reported redness or swelling, and the publication indicates that in most cases, such events resolved within 1 to 2 days.
Participants also experienced systemic effects, including fever (in most cases lower than 38.9°C and more common after dose 2), fatigue (25-50% of participants depending on age group and dose), headache (25-50% of participants depending on age group and dose), chills, and muscle or joint pain; more rarely, patients could experience gastrointestinal effects such as vomiting or diarrhea.
As with the local events, these events were almost always grade I or II.
While some events were reported by the placebo groups, these events were much rarer than in the treatment group even though compliance was similar.
Based on the efficacy and safety information released, the vaccine was approved in early December by the United Kingdom’s Medicines and Healthcare Products Regulatory Agency with administration outside of a clinical trial beginning on December 8, 2020 [928,929].
As of December 11, 2020, the United States FDA approved this vaccine under an emergency use authorization [930].
7.6 Viral Vector Vaccines
As discussed earlier, delivery and presentation of antigens are key to induce immunity against SARS-CoV2.
Viral vectors have emerged as a safe and efficient method to deliver foreign substances to induce an efficient immune response [931].
These vaccines use a different virus to deliver genetic material from the target virus of interest.
The vaccine uses the host machinery to construct antigen(s) from the transported genetic material.
There are several viral vector vaccines that are available for veterinary use [932].
While there is currently only one FDA-approved viral vector vaccine for the use in humans against Ebola [doi:10.1016/j.cell.2020.03.011], several Phase I and Phase II clinical trial are ongoing [931].
These vaccines are able to induce both an antibody and cellular response; however, the response is limited due to the immunogenicity of the viral vector used [933,934].
Various viral vector platforms including Poxviruses, Adenoviruses, and Vesicular Stomatitis Viruses (VSV) are being developed for treating numerous infectious diseases such as Malaria, Ebola, HIV, and more recently, SARS-CoV-2.
Here we review the current state of the art for developing viral vector vaccines against SARS-CoV-2.
7.6.1 Sputnik-V (Gam-COVID-Vac and Gam-COVID-Vac-Lyo)
The vaccine Gam-COVID-Vac, nicknamed Sputnik V in reference to the space race and “V for vaccine”, was developed by the Gamaleya National Center of Epidemiology and Microbiology in Moscow.
Gamaleya is an organization with prior experience using the adenovirus platform for the development of vaccines for Middle East respiratory syndrome-related coronavirus and Ebola virus, although neither of the previous vaccines were internationally licensed [935].
The development of Sputnik V was financed by the Russian Direct Investment Fund (RDIF) [936,937].
Sputnik V is a replication-deficient recombinant adenovirus (rAd) vaccine that combines two adenovirus vectors, rAd26-S and rAd5-S, that express the full-length SARS-CoV-2 spike glycoprotein.
These vectors are intramuscularly administered individually using two separate vaccines in a prime-boost regimen.
The rAd26-S is administered first, followed by rAd5-S 21 days later.
Both vaccines deliver 1011 viral particles per dose.
This approach is designed to overcome any potential pre-existing immunity to adenovirus in the population [938], as some individuals may possess immunity to Ad5 [939].
Sputnik V is the only recombinant adenovirus vaccine to utilize two vectors.
Other vaccines, such as the Oxford-AstraZeneca vaccine, utilize the chimpanzee adenovirus vector (ChAdOx1 nCoV-19) for both doses [940].
The Sputnik V vaccines are available in both a lyophilized (Gam-COVID-Vac-Lyo) and frozen form (Gam-COVID-Vac), which are stored at 2-8°C and -18°C respectively [941].
The lyophilized vaccine is convenient for distribution and storage, particularly to remote or disadvantaged areas [942].
In the race to develop vaccines against SARS-CoV-2, President Vladimir Putin of Russia announced the approval of the Sputnik V vaccines on August 11th, 2020 in the absence of clinical evidence [936].
Consequently, many international scientific agencies and public health bodies expressed concern that due diligence to the clinical trial process was subverted for the sake of expediency, leading many to question the safety and efficacy of Sputnik V [936,943,944].
Despite regulatory, safety, and efficacy concerns, pre-orders for 1 billion doses of the Sputnik V were reported within days of the vaccine’s approval in Russia [936].
Almost a month later, the phase I/II trial data was published [941].
In the phase I/II trial study conducted between late June and early August 2020, 76 participants (18-60 years old) were split into two groups of 38 participants, which were non-randomized in two hospitals in Russia.
In phase I, 9 patients received rAd26 and 9 patients received rAd5-S to assess safety over 28 days.
In phase II, at least 5 days after the completion of phase I, 20 patients received a prime-boost vaccination of rAd26-S on day 0 and rAd5-S on day 2, which was administered intramuscularly.
The phase I/II trial reported that both vaccines were deemed safe and well tolerated.
The most common adverse events reported were mild, such as pain at the injection site (58%), hypothermia (50%), headaches (42%), fatigue (28%), and joint and muscle pain (24%).
Seroconversion was observed in all participants three weeks post the second vaccination (day 42), and all participants produced antibodies to the SARS-CoV-2 glycoprotein.
RBD-specific IgG levels were high in both the frozen and lyophilized versions of the vaccine (14,703 and 11,143 respectively), indicating a sufficient immune response to both.
Three weeks post the second vaccination, the virus-neutralizing geometric mean antibody titers were 49.25 and 45.95 from the frozen and lyophilized vaccines, respectively.
At 28 days, median cell proliferation of 1.3% CD4+ and 1.1% CD8+ were reported for the lyophilized vaccine and 2.5% CD4+ and 1.3% CD8+ for the vaccine stored frozen.
These results indicated that both forms of Sputnik V appeared to be safe and induce a humoral and cellular response in human subjects [941], which may be robust enough to persist and not wane rapidly [938].
A press release on November 11th, 2020 indicated positive results from an interim analysis of the phase III Sputnik V trials, which reported 92% efficacy in 16,000 volunteers [945].
However, this release came only two days after both Pfizer and BioNTech reported that their vaccines had an efficacy over 90%, which led to significant skepticism of the Russian findings for a myriad of reasons including the lack of a published protocol and the “reckless” approval of the vaccine in Russia months prior to the publication of the interim results of the phase III trial [945,946].
In February 2021, the interim results of the phase III randomized, double-blind, placebo-controlled trial were eventually published in The Lancet[947].
The multicenter trial enrolled 21,862 participants over the age of 18 years old in Moscow, Russia.
The participants were randomly assigned to receive either a 0.5 mL/dose of vaccine or placebo, which was comprised of the vaccine buffer composition, that was delivered intramuscularly using the same prime-boost regimen as in the phase I/II trials.
From September 7th to Nov 24th, 19,866 participants completed the trial.
Of the 14,964 participants who received the vaccine, 16 (0.1%) were confirmed to have COVID-19, whereas 62 of the 4,902 participants (1.3%) in the placebo group were confirmed to have COVID-19.
Of these participants, no moderate or severe cases of COVID-19 were reported in the vaccine group, juxtaposed with 20 in the placebo group.
However, only symptomatic individuals were confirmed for SARS-CoV-2 infection in this trial.
Therefore, asymptomatic infections were not detected, thus potentially inflating the efficacy estimate.
Overall, a vaccine efficacy of 91.6% (95% CI 85.6-95.2) was reported, where an efficacy of 91.8% was reported for those over 60 years old and 92.7% for those who were 51-60 years old.
Indeed, 14 days after the first dose, 87.6% efficacy was achieved and the immunity required to prevent disease occurred within 18 days of vaccination.
Based on these results, scientists are investigating the potential for a single dose regimen of the rAd26-S sputnik V vaccine [948].
By the end of the trial, 7,485 participants reported adverse events, of which 94% were grade I.
Of the 68 participants who experienced serious adverse events during the trial, 45 from the vaccine group and 23 from the placebo groups, none were reported to be associated with the vaccination.
Likewise, 4 deaths occurred during the trial period that were not related to the vaccine [947].
The interim findings of the phase III trial indicate that the Sputnik V vaccine regimen appears to be safe with 91.6% efficacy.
Gamaleya had intended to reach a total of 40,000 participants for the completion of their phase III trial.
However, the trial has stopped enrolling participants and the numbers have been cut to 31,000 as many individuals in the placebo group dropped out of the study to obtain the vaccine [949].
Indeed, other trials involving Sputnik V are currently underway in Belarus, India, the United Arab Emirates, and Venezuela [950].
Preliminary results of a trial of Argentinian healthcare workers in Buenos Aires who were vaccinated with the Sputnik V rAd26-R vector-based vaccine seems to support the short term safety of the first vaccination [951].
Of the 707 vaccinated healthcare workers, 71.3% of the 96.6% of respondents reported at least one adverse event attributed to the vaccine.
Of these individuals, 68% experienced joint and muscle pain, 54% had injection site pain, 11% reported redness and swelling, 40% had a fever, and 5% reported diarrhea.
Only 5% of the vaccinated participants experienced serious adverse events that required medical attention, of which one was monitored as an inpatient.
Additionally, an Independent assessment of Sputnik V in a phase II clinical trial in India found the vaccine to be effective, but the data is not yet publicly available [952].
On December 21st, 2020, Gamaleya, AstraZeneca, R-Pharm, and the Russian Direct Investment Fund agreed to assess the safety and immunogenicity of the combined use of components of the AstraZeneca and University of Oxford AZD1222 (ChAdOx1) vaccine and the rAd26-S component of the Sputnik V vaccine in clinical trials [953].
This agreement hopes to establish scientific and business relations between the entities with an aim to co-develop a vaccine providing long-term immunization.
The trial, which will begin enrollment soon, will include 100 participants in a phase II open-label study and is hoped to be complete within 6 months.
Participants will first receive an intramuscular dose of AZD1222 on day 1, followed by a dose of rAd26 on day 29.
Participants will be monitored from day 1 for 180 days in total.
The primary outcomes measured will include incidence of serious adverse events post first dose until the end of the study.
Secondary outcome measures will include incidence of local and systemic adverse events 7 days post each dose, a time course of antibody responses for the Spike protein and the presence of anti-SARS-CoV-2 neutralizing antibodies [954].
Overall, there is hesitancy surrounding the management of the Sputnik V vaccine approval process and concerns over whether the efficacy data may be inflated due to a lack of asymptomatic testing within the trial.
However, the interim results of the phase III study were promising and further trials are underway, which will likely shed light on the overall efficacy and safety of the Sputnik V vaccine regimen.
There may be some advantage to the Sputnik V approach including the favorable storage conditions afforded by choice between a frozen and lyophilized vaccine.
Furthermore, the producers of Gam-COVID-Vac state that they can produce the vaccine at a cost of less than $10 per dose or less than $20 per patient [955].
7.6.2 Janssen’s JNJ-78436735
The Johnson & Johnson (J&J) vaccine developed by Janssen Pharmaceuticals, Inc., a subsidiary of J&J, was conducted in collaboration with and funded by “Operation Warp Speed” [956,957].
The vaccine candidate JNJ-78436735, formerly known as Ad26.COV2-S, is a monovalent vaccine that is composed of a replication-deficient adenovirus serotype 26 (Ad26) vector expressing the stabilized pre-fusion S protein of SARS-CoV-2 [958,959].
The vaccine was developed using Janssen’s AdVac® and PER.C6 platforms that were previously utilized to develop the European Commission-approved Ebola vaccine (Ad26 ZEBOV and MVN-BN-Filo) and their Zika, respiratory syncytial (RSV), and human immunodeficiency virus (HIV) investigational vaccine candidates [960].
The development of a single-dose vaccine was desirable by J&J from the outset, with global deployment being a key priority [961].
Using their AdVac® technology, the vaccine can remain stable for up to two years between -15℃ and -25℃ and at least three months at 2-8℃ [960].
This allows the vaccine to be distributed easily without the requirement for very low temperature storage, unlike many of the other COVID-19 vaccine candidates.
J&J screened numerous potential vaccine candidates in vitro and in animal models using varying different designs of the S protein, heterologous signal peptides, and prefusion-stabilizing substitutions [959].
A select few candidates were further investigated as a single dose regimen in Syrian golden hamsters, a single dose regimen in rhesus macaques, and a single- and two-dose regimen in both adult and aged rhesus macaques [959,961,962,963].
From these studies, the JNJ-78436735 candidate was selected for its favorable immunogenicity profile and ease of manufacturability [959,961,962,963].
A SARS-CoV-2 challenge study in rhesus macaques showed that vaccine doses as low as 2 x 109 viral particles/mL was sufficient to induce strong protection in bronchoalveolar lavage but that doses higher than 1.125 x 1010 were required to close achieve close to complete protection in nasal swabs [964].
Indeed, six months post-immunization, levels of S-binding and neutralizing antibodies in rhesus macaques indicated that the JNJ-78436735 vaccine conferred durable protection against SARS-CoV-2 [965].
Following selection of the JNJ-78436735 vaccine, J&J began phase I/IIa trials.
The interim phase I/IIa data was placed on the medRxiv preprint server on September 25th, 2020 [966] and was later published in the New England Journal of Medicine on January 13th, 2021 [958].
The phase I/IIa multi-center, randomized, placebo-controlled trial enrolled 402 healthy participants between 18-55 years old and a further 403 healthy older participants ≥ 65 years old [958].
Patients were administered either a placebo, a low dose (5 x 1010 viral particles per mL), or a high dose (1 X 1011 viral particles per mL) intramuscularly as part of either a single- or two-dose regimen.
All patients received injections 56 days apart, but participants in the single-dose condition received the placebo at the second appointment.
Those who received only one dose of either vaccine received a placebo dose at their second vaccination visit.
A comparison of the single versus double dose regimen has yet to be published.
The primary endpoints of both the trial were safety and reactogenicity of each dose.
Fatigue, headache, myalgia, and pain at the injection site were the most frequent solicited adverse events reported by participants.
Although less common, particularly for those in the elderly cohort and those on the low dose regimen, the most frequent systemic adverse effect was fever.
Overall, immunization was well tolerated, particularly at the lower dose concentration.
In terms of reactogenicity, over 90% of those who received either the low or high dose demonstrated seroconversion in a neutralization assay using wild-type SARS-CoV-2, 29 days after immunization [958].
Neutralizing geometric mean ratio of antibody titers (GMT) between 224-354 were detected regardless of age.
By day 57, 100% of the 18-55 year old participants had neutralizing GMT (288-488), which remained stable until day 71.
In the ≥ 65 years old cohort, the incidence of seroconversion for the low- and high-dose was 96% and 88% respectively by day 29.
GMTs for the low and high doses were slightly lower for participants ≥ 65 years old (196 and 127 respectively), potentially indicating slightly lower immunogenicity.
Seroconversion of the S antibodies was detected in 99% of individuals between 18-55 years old for the low and high doses (GMTs 528 and 695 respectively), with similar findings reported for the ≥ 65 years old.
Indeed, both dose concentrations also induced robust Th1 cytokine-producing S-specific CD4+ T cells and CD8+ T cell responses in both age groups.
The findings of the phase I/IIa study supported further investigation of a single immunization using the low dose vaccine.
Therefore, 25 patients were enrolled for a second randomized double-blind, placebo-controlled phase 1 clinical trial currently being conducted in Boston, Massachusetts for 2 years [967].
Participants received either a single dose followed by a placebo, or a double dose of either a low dose (5 x 1010 viral particles/mL) or a high dose (1 x 1011 viral particles/mL) vaccine administered intramuscularly on day 1 or day 57.
Placebo-only recipients received a placebo dose on day 1 and 57.
Interim analyses conducted on day 71 indicated that binding and neutralizing antibodies developed 8 days after administration in 90% and 25% of vaccine recipients, respectively.
Binding and neutralizing antibodies were detected in 100% of vaccine recipients by day 57 after a single dose immunization.
Spike-specific antibodies were highly prevalent (GMT 2432 to 5729) as were neutralizing antibodies (GMT 242 to 449) in the vaccinated groups.
Indeed, CD4+ and CD8+ T-cell responses were also induced, which may provide additional protection, particularly if antibodies wane or poorly respond to infection [968].
On September 23rd, 2020, J&J launched its phase III trial ENSEMBLE and released the study protocol to the public [960,969].
The trial intended to enroll 60,000 volunteers to assess the safety and efficacy of the single vaccine dose versus placebo with primary endpoints of 14 and 28 days post-immunization [960].
The trial was conducted in Argentina, Brazil, Chile, Colombia, Mexico, Peru, South Africa, and the U.S.
The trial was paused briefly in October 2020 to investigate a “serious medical event”, but resumed shortly after [970].
An interim analysis was reported via press release on January 29th, 2021 [971,972].
The interim data included 43,783 participants who accrued 468 symptomatic cases of COVID-19.
It was reported that the JNJ-78436735 vaccine was 66% effective across all regions studied for the prevention of moderate to severe COVID-19 28 days post-vaccination in those aged 18 years and older.
Notably, JNJ-78436735 was 85% effective for the prevention of laboratory-confirmed severe COVID-19 and 100% protection against COVID-19-related hospitalization and death 28 days post-vaccination across all study sites.
Efficacy of the vaccine against severe COVID-19 increased over time, and there were no cases of COVID-19 reported in immunized participants after day 49.
The trial also determined that the vaccine candidate has a favorable safety profile as determined by an independent Data and Safety Monitoring Board.
The vaccine was well tolerated, consistent with previous vaccines produced using the AdVac® platform.
Fever occurred in 9% of vaccine recipients, with grade 3 fever occurring in only 0.2% of recipients.
Serious adverse events were reportedly higher in the placebo group than the vaccine group, and no anaphylaxis was reported [972].
At the time the phase III trial was being conducted, several concerning variants, including B.1.1.7 [277] and B.1.351 [282], were spreading across the globe.
In particular, B.1.351 was first identified in South Africa, which was one of the JNJ-78436735 vaccine trial sites.
Therefore, the J&J investigators also analyzed the efficacy of the JNJ-78436735 vaccine associated with their various trial sites to determine any potential risk of reduced efficacy as a result of the novel variants.
It was determined that JNJ-78436735 was 72% effective in the U.S., 66% effective in Latin America, and 57% effective in South Africa 28 days post-vaccination.
These findings underpin the importance of monitoring for the emergence of novel SARS-CoV-2 variants and determining their effects on vaccine efficacy.
Looking forward, Janssen are also running a phase III randomized, double-blind, placebo-controlled clinical trial, Ensemble 2, which aims to assess the efficacy, safety, and immunogenicity of a two-dose regimen of JNJ-78436735 administered 57 days apart.
This trial will enroll 30,000 participants ≥ 18 years old from Belgium, Colombia, France, Germany, Philippines, South Africa, Spain, U.K., and the U.S. [973].
This trial will also include participants with and without comorbidities associated with an increased risk of COVID-19.
7.7 Inactivated Whole Virus Vaccines
These types of vaccines use full virus particles that have been rendered non-infectious by chemical (i.e. using formaldehyde or β-propiolactone [974]) or physical means (i.e. heat).
Though these virus particles are inactivated they still have the capacity to prime the immune system.
The size of the virus particle makes it ideal for uptake by antigen presenting cells (APC), which leads to stimulation of helper T-cells [975].
Additionally, the array of epitopes on the surface of the virus increases antibody binding efficiency [975].
The native conformation of the surface proteins, which is also important for eliciting an immune response, is preserved using these techniques.
Membrane proteins, which support B-cell response to surface proteins, are also included using this method [976].
Overall, these vaccines are able to mimic the key properties of the virus that stimulate a robust immune response, but the risk of adverse reactions is reduced because the virus is inactivated, thus unable to replicate.
Some common successful whole virus vaccine include targeting influenza viruses, poliovirus, and hepatitis A virus.
7.7.1 Sinovac’s CoronaVac
The CoronaVac vaccine is being developed by Sinovac, a Beijing-based biopharmaceutical company.
The vaccine is using an inactivate whole virus with the addition of an aluminum adjuvant [977].
The vaccine is currently in Phase III clinical trials.
Pre-clinical trials were performed using BALB/c mice and rhesus macaques [978].
The SARS-CoV-2 strains used in this trial isolated from 11 hospitalized patients (5 from China, 3 from Italy, 1 from the UK, 1 from Spain, 1 from Switzerland).
A phylogenetic analysis demonstrated that the strains were representative of the current circulating variants.
One of the strains, CN2, from China was used as the inactivated and purified virus while the other 10 strains were used to challenge.
The CN2 was grown in Vero cells.
An ELISA assay was used to assess the immunogenicity of the vaccine.
10 mice were injected with the vaccine on day 0 and day 7 with varying doses (0, 1.5, 3 or 6 μg), and 10 mice were treated with physiological saline as the control.
IgG developed in the serum of all vaccinated mice.
Using the same setup, immunogenicity was also assessed in macaques.
Four macaques were assigned to each of four groups: treatment with 3 μg at day 0, 7 and 14, treatment with a high dose of 6 μg at day 0, 7 and 14, administration of a placebo vaccine, and administration of only the adjuvant.
All vaccinated macaques induced IgGs and neutralizing antibodies.
After challenge with SARS-CoV-2 strain CN1, vaccinated macaques were protected compared to control macaques (placebo or adjuvant only) based on histology and viral loads collected from different regions of the lung.
Phase I/II clinical trials were conducted in adults 18-59 years old [979] and adults over 60 years old [980] in China.
In the case of adults 18-59 years old, a single center, randomized, double-blind, placebo-controlled phase I/II trial was conducted in April 2020.
Patients in this study were recruited from the community in Suining County of Jiangsu province, China.
For the phase I trial, 144 (of 185 screened) participants were enrolled, with 72 enrolled in the 14-day interval cohort (i.e., treated on day 0 and day 14) and 72 in the 28-day interval cohort.
This group of 72 participants was split into 2 blocks for a low-dose (3 μg) and high-dose (6 μg) vaccine.
Within each block, participants were randomly assigned vaccination with CoronaVac or placebo (aluminum diluent without the virus) at a 2:1 ratio.
Both the vaccine and placebo were prepared in a Good Manufacturing Practice-accredited facility of Sinovac Life Sciences (Beijing, China).
The phase II trial followed the same organization of participants, this time using 300 enrolled participants in the 14-day and another 300 enrolled in the 28-day groups.
One change of note was that the vaccine was produced using a highly automated bioreactor (ReadyToProcess WAVE 25, GE, Umeå, Sweden) to increase vaccine production capacity.
This change resulted in a higher intact spike protein content.
The authors of this study were not aware of this antigen-level difference between the vaccine batches for the phase I/II when the ethical approval for the trials occurred.
To assess adverse responses, participants were asked to record any events up to 7 days post-treatment.
The reported adverse events were graded according to the China National Medical Products Administration guidelines.
In the phase I trial, the overall incidence of adverse reactions was 29-38% of patients in the 0 to 14 day group and 13-17% in the 0 to 28 day vaccination group.
The most common symptom was pain at the injection site, which was reported by 17-21% of patients in the 0 to 14 day cohort and 13% in the 0 to 28 day cohort.
Most adverse reactions were mild (grade 1) where patients recovered within 48 hours.
A single case of acute hypersensitivity with manifestation of urticaria 48 hours following the first dose of study drug was reported in the 6 μg group
Most adverse reactions were mild (grade 1) in severity and participants recovered within 48 hours.
There was a single case, from the 6 μg group, of acute hypersensitivity with manifestation of urticaria 48 hours after the first dose.
Both the 14-day and 28-day cohorts had a strong neutralizing Ab response.
The neutralizing Ab response was measured using a micro cytopathogenic effect assay, which assesses the minimum dilution of neutralizing Ab to be 50% protective against structural changes in host cells in response to viral infection [981].
Additionally IgG antibody titers against the receptor binding domain were also measured using ELISA.
Another phase I/II study was performed with older patients (older than 60 years) [980].
The study conducted a single-center, randomized, double-blind, placebo-controlled trial.
The phase I trial looked at dose escalation using 3 dosages: 1.5, 3 and 6 μg.
The mean age of participants was 65.8 years (std = 4.8).
Of 95 screened participants, 72 were enrolled.
These 72 participants were split into low (3 μg) and high (6 μg) dose groups.
Within each group, 24 participants received the treatment and 12 the placebo.
A neutralizing antibody response against live SARS-CoV-2 was detected compared to baseline using the same micro cytopathogenic effect assay.
This response was similar across the two dose concentrations.
Additionally, they did not observe a difference in response between age groups (60–64 years, 65–69 years, and ≥70 years).
In phase II the mean age was 66.6 years (standard deviation = 4.7).
499 participants were screened and 350 were enrolled.
300 were evenly split into 1.5, 3 and 6 μg dose groups, and the remaining 50 were assigned to the placebo group.
Again, they found a neutralizing antibody response in phase II.
There wasn’t a significant different between the response to 3 μg versus 6 μg, but the response was higher than that to 1.5 μg.
Participants were required to record adverse reaction events within the first 7 days after each dose.
The safety results were combined across phage I and II.
All adverse reactions were either mild (grade 1) or moderate (grade 2) in severity.
The most common symptom was pain at the injection site (9%) and fever (3%).
2% (7 participants) reported severe adverse events (4 from the 1.5 μg group, 1 from the 3 μg group, 2 from the 6 μg group), though these were found to be unrelated to the vaccine.
Overall, the results from the pre-clinical and phase I/II clinical trials are promising.
It was also very hopeful to see that the immune response was consistent in older adults (> 60 years).
Currently, phase III trials are being conducted in Brazil [982].
This is a randomized, multicenter, endpoint driven, double-blind, placebo-controlled clinical trial.
They are expecting 13,060 participants with 11,800 ages 18 to 59 years and 1,260 age 60+.
Participants will be health care professionals.
7.8 Protein Subunit Vaccines
Compared to the inactivated whole virus vaccines, these protein subunit vaccines isolate a single protein of the virus and use it to stimulate the immune system.
These proteins, also referred to as antigens, are usually those located on the surface of the viral particle and are therefore key targets of the immune system.
These proteins are typically grown in yeast and then harvested.
This vaccine can stimulate antibodies and CD4+ T-cell response [983].
The main advantage of this method is that they are considered very safe because the antigen alone cannot cause an infection; however, the immune response is weaker and an adjuvant is usually needed to boost the response [984].
7.8.1 Novavax NVX-CoV2373
Novavax-CoV2373 is a protein nanoparticle vaccine candidate against SARS-CoV-2.
The vaccine is constructed from a mutated SARS-CoV-2 spike protein in combination with a specialized adjuvant to elicit an immune response against SARS-CoV-2.
The spike protein is recombinantly expressed in Sf9 insect cells [985], which have previously been used for several other FDA-approved protein therapeutics [986].
The expressed spike protein contains mutations in the furin cleavage site (682-RRAR-685 to 682-QQAQ-685) to avoid cleavage of the spike protein as well as two proline substitutions (K986P and V987P) to improve thermostability [985].
The improved stability caused by the proline substitutions is particularly critical to facilitating global distribution, particularly to regions where local refrigerator/freezer capacities are limited.
Importantly, these amino acid substitutions did not affect the ability of the spike protein to bind the hACE2 receptor (the target receptor of SARS-CoV-2 spike protein).
The Novavax-CoV2373 vaccine candidate uses a proprietary, saponin-based Matrix-MTM adjuvant that contains two different 40nm-sized particles formed by formulating purified saponin with cholesterol and phospholipids [987].
In preclinical models, the use of the Matrix-M adjuvant potentiated the cellular and humoral immune responses to influenza vaccines [???- 2659.2011.00256.x,987,988,989].
Importantly, Matrix-M adjuvant-containing vaccines have shown acceptable safety profiles in human clinical trials [??? 042].
In preclinical mouse models, Novavax-CoV2373 elicited high anti-spike IgG titers 21-28 days post-vaccination that could neutralize the SARS-CoV-2 virus and protect the animals against virus challenge [985].
Antibody titers were significantly elevated in groups receiving the vaccine with the Matrix-M adjuvant compared to the groups without adjuvant.
Novavax-CoV2373 was able to induce a multifunctional CD4/CD8 T-cell responses and generate high frequencies of follicular helper T-cells and B-cell germinal centers after vaccination.
These findings were subsequently evaluated in a baboon primate model, in which Novavax-CoV2373 also elicited high antibody titers against the SARS-CoV-2 spike protein, as well as an antigen specific T-cell response.
Based on this data Novavax initiated a Phase 1/2 clinical trial to evaluate the safety and immunogenicity of Novavax-CoV2373 with Matrix-M (NCT04368988)[990].
The phase I/II trial was a randomized, placebo-controlled study with 131 healthy adult participants in 5 treatment arms [990].
Participants that received the recombinant SARS-CoV-2 vaccine with or without the Matrix-M adjuvant got two injections, 21 days apart.
Primary outcomes that were assessed include reactogenicity, lab-values (serum chemistry and hematology), and anti-spike IgG levels.
Secondary outcomes measured included virus neutralization, T-cell responses, and unsolicited adverse events.
The authors reported that no serious treatment-related adverse events occurred in any of the treatment arms.
Reactogenicity was mostly absent and of short duration.
The two-dose vaccine regimen induced anti-spike IgG levels and neutralizing antibody-titers exceeding those in the convalescent plasma of symptomatic patients.
In line with the preclinical studies, the use of Matrix-M adjuvant further increased anti-spike immunoglobulin levels and induced a Th1 response.
The outcomes of this trial suggest that Novavax-CoV2373 has an acceptable safety profile and is able to induce a strong immune response with high neutralizing antibody titers.
The phase II component of this phase I/II trial was recently uploaded to an open-access repository [991].
This part of the trial was designed to identify which dosing regimen should move forward into late phase clinical trials.
Both younger (18-59 years) and older patients (60-84 years) were randomly assigned to receive either 5 μg or 25 μg Novavax-CoV2373 or placebo in two doses, 21 days apart.
In line with the phase I data, reactogenicity remained mild to moderate and of short duration.
Both dose levels were able to induce high anti-spike IgG titers as well as neutralizing antibody responses after the second dose.
Based on both safety and efficacy data, the 5 μg dosing regimen was selected as the optimal dose regiment for the ongoing phase III trial.
Although the phase III trial data has not been published yet, Novavax announced an efficacy of 89.3% based on their phase 3 trial in the UK and South Africa.
This trial included over 15,000 participants in the UK and 4,000 participants in South Africa with occurrence of a PCR-confirmed symptomatic case as the primary endpoint.
In the first interim analysis (UK), 56 cases of COVID-19 were observed in the placebo group compared to 6 cases in the treatment group.
Importantly, the vaccine candidate also shows significant clinical efficacy against the prevalent UK and South African variants.
The company has also initiated the development of new constructs to select candidates that can be used as a booster against new strains and plans to initiate clinical trials for these new constructs in the second quarter of 2021.
7.9 Vaccine Development Summary
7.10 Complementary Approaches to Vaccine Development
7.10.1 Adjuvants for Vaccines
Adjuvants include a variety of molecules or larger microbial-related products that have an effect on the immune system or an immune response of interest.
They can either be comprised of or contain immunostimulants or immunomodulators.
Adjuvants are sometimes included within vaccines, especially vaccines other than live-attenuated and inactivated viruses, in order to enhance the immune response.
A review on the development of SARS-CoV-2 vaccines [992] also included a brief summary of the potential of adjuvants for these vaccines, including a brief description of some already commonly used adjuvants.
Different adjuvants can regulate different types of immune responses, so the type or combination of adjuvants used in a vaccine will depend on both the type of vaccine and concern related to efficacy and safety.
A variety of possible mechanisms for adjuvants have been researched [993,994,995], including the following: induction of DAMPs that can be recognized by certain PRRs of the innate immune system; functioning as PAMP that can also be recognized by certain PRRs; and more generally enhancing the humoral or cellular immune responses.
Selection of one or more adjuvants requires considering how to promote the advantageous effects of the components and/or immune response and, likewise, to inhibit possible deleterious effects.
There are also considerations related to the method of delivering (or co-delivering) the adjuvant and antigen components of a vaccine.
7.10.2 Trained Immunity
Another approach that is being investigated explores the potential for vaccines that are not made from the SARS-CoV-2 virus to confer what has been termed trained immunity.
In a recent review [996], trained immunity was defined as forms of memory that are temporary (e.g., months or years) and reversible.
It is induced by exposure to whole-microorganism vaccines or other microbial stimuli that generates heterologous protective effects.
Trained immunity can be displayed by innate immune cells or innate immune features of other cells, and it is characterized by alterations to immune responsiveness to future immune challenges due to epigenetic and metabolic mechanisms.
These alterations can take the form of either an increased or decreased response to immune challenge by a pathogen.
Trained immunity elicited by non-SARS-CoV-2 whole-microorganism vaccines could potentially improve SARS-CoV-2 susceptibility or severity [997].
One type of stimulus which research indicates can induce trained immunity is bacillus Calmette-Guerin (BCG) vaccination.
BCG is an attenuated form of bacteria Mycobacterium bovis.
The vaccine is most commonly administered for the prevention of tuberculosis in humans.
Clinical trials in non-SARS-CoV-2-infected adults have been designed to assess whether BCG vaccination could have prophylactic effects against SARS-CoV-2 by reducing susceptibility, preventing infection, or reducing disease severity.
A number of trials are now evaluating the effects of the BCG vaccine or the related vaccine VPM1002 [997,998,999,1000,1001,1002,1003,1004,1005,1006,1007,1008,1009,1010,1011].
The ongoing trials are using a number of different approaches.
Some trials enroll healthcare workers, other trials hospitalized elderly adults without immunosuppression who get vaccinated with placebo or BCG at hospital discharge, and yet another set of trials older adults (>50 years) under chronic care for conditions like hypertension and diabetes.
One set of trials, for example, uses time until first infection as the primary study endpoint; more generally, outcomes measured in some of these trials are related to incidence of disease and disease severity or symptoms.
Some analyses have suggested a possible correlation at the country level between the frequency of BCG vaccination (or BCG vaccination policies) and the severity of COVID-19 [997].
Currently it is unclear whether this correlation has any connection to trained immunity.
Many possible confounding factors are also likely to vary among countries, such as age distribution, detection efficiency, stochastic epidemic dynamic effects, differences in healthcare capacity over time in relation to epidemic dynamics, and these have not been adequately accounted for in current analyses.
It is unclear whether there is an effect of the timing of BCG vaccination, both during an individual’s life cycle and relative to the COVID-19 pandemic.
Additionally, given that severe SARS-CoV-2 may be associated with a dysregulated immune response, it is unclear what alterations to the immune response would be most likely to be protective versus pathogenic (e.g., [146,997,1012,1013]).
The article [997] proposes that trained immunity might lead to an earlier and stronger response, which could in turn reduce viremia and the risk of later, detrimental immunopathology.
While trained immunity is an interesting possible avenue to complement vaccine development efforts through the use of an existing vaccine, additional research is required to assess whether the BCG vaccine is likely to confer trained immunity in the case of SARS-CoV-2.
7.11 Viral evolution and vaccine protection
With these vaccines in place, one concern is how the virus’s continued evolution will affect their efficacy.
Since the start of this pandemic, we have already seen multiple variants emerge: B.1.1.7, which emerged in the UK, B.1.351, which emerged in South Africa, and P.1, which emerged in Brazil.
Viruses evolve or mutate at different rates.
Mutation rate is measured as the number of substitutions per nucleotide per cell infected (μs/n/c) [1014].
RNA viruses tend to have mutation rates between 10-6 to 10-4[1014].
As a reference, influenza A virus has a mutation rate of 10-5, whereas the mutation rate of SARS-CoV-2 is lower, with the mutation rate estimated at 10-6[1015].
The accumulation of mutations allows the virus to escape recognition by the immune system [1016].
The efficacy of vaccines depends on their ability to train the immune system to recognize the virus.
Therefore, viruses can develop resistance to vaccines through the accumulation of mutations that affect recognition.
The lower mutation rate of SARS-CoV-2 suggests the possibility of SARS-CoV-2 vaccines having a more long-lasting effect compared to vaccines targeting the influenza A virus.
The current SARS-CoV-2 vaccines in distribution have been reported to provide similar efficacy against the B.1.1.7 variant compared to the variants common at the time they were developed but reduced efficacy against the B.1.351 variant [1017].
Pfizer and Moderna announced that they are working on developing a booster shot to improve efficacy against the B.1.351 variant [1018].
The WHO continues to monitor the emergence of variants and their impact on vaccine efficacy [1019].
Previous research in the computational prediction of the efficacy of vaccines targeting the influenza A virus might complement efforts to monitor these types of viral outbreaks [1020].
To adapt, future vaccines may need to account for multiple variants and strains of SARS-CoV-2, and booster shots may be required [1021].
7.12 Global Vaccine Status and Distribution
The unprecedented development of COVID-19 vaccines in under a year since the beginning of the pandemic now requires rapid global vaccine production and distribution plans.
The development of vaccines is costly and complicated, but vaccine distribution can be just as challenging.
Logistical considerations such as transport, storage, equipment (e.g., syringes), the workforce to administer the vaccines, and a continual supply from the manufacturers to meet global demands all must be accounted for and will vary globally due to economic, geographic, and sociopolitical reasons [1022,1023,1024].
Deciding on the prioritization and allocation of the COVID-19 vaccines is also a challenging task due to ethical and operational considerations.
Various frameworks, models, and methods have been proposed to tackle these issues with many countries, regions or states as is the case in the U.S., devising their own distribution and administration plans [1025,1026,1027,1028,1029].
The majority of the distribution plans prioritize offering vaccines to key workers such as health care workers, and those who are clinically vulnerable such as the elderly, the immunocompromised, and individuals with comorbidities, before targeting the rest of the population, who are less likely to experience severe outcomes from COVID-19 [1030].
As of March 6th, 2021, approximately 319 million vaccine doses have been administered in at least 118 countries worldwide using 10 different vaccines [1031,1032].
The global vaccination rate is currently ~8.1 million doses per day, which at the current rate would take almost 4 years to vaccinate 75% of the world’s population according to media estimates of a two-dose regimen [1032].
Vaccine production and distribution varies from region to region and seems to depend on the availability of the vaccines and potentially a country’s resources and wealth [1033].
In North America, the majority of vaccines distributed until March 2021 have been produced by Pfizer-BioNTech and Moderna.
In Canada, the vaccine approval process is conducted by Health Canada, which uses a fast-tracked process whereby vaccine producers can submit data as it becomes available to allow for rapid review.
An approval may be granted following reviews of the available phase III clinical data.
This is followed by a period of pharmacovigilance in the population using their post-market surveillance system, which will monitor the long-term safety and efficacy of any vaccines [1034,1035].
Health Canada has authorized the use of the Pfizer (December 9th, 2020), Moderna (December 23rd, 2020), Oxford-AstraZeneca (February 26th, 2021), and the Janssen (March 5th, 2021) vaccines, and the Novavax Inc vaccine is also under consideration [1036].
While Canada initially projected that by the end of September 2021 a vaccine would be available for all Canadian adults, they now predict that it may be possible earlier as more vaccines have been approved and become available [1037].
In the U.S., vaccines are required to have demonstrated safety and efficacy in phase III trials before manufacturers apply for an emergency use authorization (EUA) from the FDA.
If an EUA is granted, an additional evaluation of the safety and efficacy of the vaccines is conducted by the CDC’s Advisory Committee on Immunization Practices (ACIP) who also provide guidance on vaccine prioritization.
On December 1st, 2020, ACIP provided an interim phase 1a recommendation that healthcare workers and long-term care facility residents should be the first to be offered any vaccine approved [1038].
This was shortly followed by an EUA on December 11th, 2020 for the use of the Pfizer-BioNTech COVID vaccine [1039], which was distributed and administered to the first healthcare workers on December 14th, 2020 [1040].
Shortly thereafter, an EUA for the Moderna vaccine was issue on December 18th, 2020 [1041].
On December 20th, 2020, ACIP updated their initial recommendations to suggest that vaccinations should be offered to people aged 75 years old and older and to non-healthcare frontline workers in phase 1b [1042].
On the same date, it was recommended that phase 1c should include people aged 65-74 years old, individuals between the ages of 16-74 years old at high-risk due to health conditions, and essential workers ineligible in phase 1b [1042].
On the following day, December 21st, 2020, the first Moderna vaccines used outside of clinical trials were administered to American healthcare workers, which was the same day that President-elect Biden and Dr. Biden received their first doses of the Pfizer-BioNTech vaccine live on television to instill confidence in the approval and vaccination process [1043].
On February 27th, 2020, the FDA issued an EUA for the Janssen COVID-19 Vaccine [1044].
This was followed by an update on recommendations by ACIP for the use of the Janssen COVID-19 vaccine for those over 18 years old [1045].
The Janssen vaccine was first distributed to healthcare facilities on March 1st, 2021.
On March 12, 2021, the WHO added the Janssen vaccine to the list of safe and effective emergency tools for COVID-19 [1046].
While the CDC’s ACIP can provide recommendations, it is up to the public health authorities of each state, territory, and tribe to interpret the guidance and determine who will be vaccinated first [1047].
Prior to distribution of the Janssen vaccine, over 103 million doses of the Moderna and Pfizer-BioNTech vaccines were delivered across the U.S., with almost 79 million doses administered.
Of the total population, 15.6% have received at least one dose and 7.9% have received a second dose of either the Moderna (~38.3 million) or the Pfizer-BioNTech (~40.2 million) vaccines by February 28th, 2021 [1048].
President Biden’s administration has predicted that by the end of May 2021 there may be enough vaccine supply available for all adults in the U.S. [1049,1050].
However, vaccine production, approval, and distribution was not straightforward in the U.S., as information was initially sparse and the rollout of vaccines was complicated by poor planning and leadership due to political activities prior to the change of administration in January 2021 [1051].
These political complications highlight the importance of the transparent vaccine approval process conducted by the FDA [1052].
Outside the U.S., the Moderna and Pfizer-BioNTech vaccines have been administered in 29 and 69 other countries, respectively, mainly in Europe and North America [1031].
The Janssen vaccine has so far only been administered in South Africa and the U.S. [1031; https://www.biopharma-reporter.com/Article/2021/02/18/South-Africa-starts-administering-Janssen-COVID-19-vaccine-to-health-workers], but it has also been approved in Bahrain, the European Union (E.U.), Iceland, Liechtenstein, and Norway[1053].
On March 11th, 2021, Johnson & Johnson received approval from the European Medicines Agency (EMA) for conditional marketing authorization of their vaccine [1054].
Notably, on March 2nd, 2021, rivals Johnson & Johnson and Merck announced that they entered an agreement to increase production of the Janssen vaccine to meet global demand [1055].
The U.K. was the first country to approve use of the Pfizer-BioNTech vaccine on December 2nd, 2020 [1056], and it was later approved by EMA on December 21st, 2020 [1057].
The U.K. was also the first to administer the Pfizer-BioNTech vaccine, making it the first COVID-19 vaccine supported by phase III data to be administered outside of clinical trials on December 8th, 2020.
The Oxford-AstraZeneca vaccine, was approved by the Medicines and Healthcare Products Regulatory Agency (MHRA) in the U.K. and by EMA in the E.U. on December 30th (2020) [1058] and January 29th (2021) [1059] respectively.
The Oxford-AstraZeneca vaccine was first administered in the UK on January 4th, 2021 [1060], and it is now being used in 53 countries in total, including Brazil, India, Pakistan, Mexico, and spanning most of Europe [1031].
The Moderna vaccine was authorized for use in the E.U. by EMA on January 6th, 2021 [1061] and in the U.K. by MHRA on January 8th, 2021 [https://www.gov.uk/government/publications/regulatory-approval-of-covid-19-vaccine-moderna].
As of March 5th, 2021, 22 million people in the U.K. had received at least one vaccine dose [1062].
While the Pfizer-BioNTech vaccine was the first to be distributed following phase III clinical trials, the first COVID-19 vaccine to be widely administered to people prior to the completion of phase III clinical trials was Sputnik V.
Sputnik V was administered to as many as 1.5 million Russians by early January [1063] due to the establishment of mass vaccination clinics in December 2020, prior to which only approximately 100,000 Russians had already been vaccinated [1064,1065].
Doses of Sputnik V have also been distributed to other parts of Europe, such as Belarus, Bosnia-Herzegovina, Hungary, San Marino, Serbia, and Slovakia [1066,1067,1068], with the Czech Republic and Austria also having expressed interest in its procurement [1069].
Hungary was the first E.U. member country to approve and distribute Sputnik V outside of Russia [1069], despite the EMA stating that they had neither approved nor received a request for approval of Sputnik V [1070].
Hungary is also in talks with China to procure the Sinopharm vaccines, which have been approved by Hungarian health authorities but also have not received approval by EMA in the E.U. [1069].
In Latin America, production facilities in both Brazil and Argentina will allow for increased production capacity of Sputnik V and doses have been distributed to Mexico, Argentina, Bolivia, Nicaragua, Paraguay, and Venezuela [1071].
Guinea was the first African nation to administer Sputnik V in December 2020, and the Central African Republic, Zimbabwe, and the Ivory Coast have all registered their interest in purchasing doses of the vaccine [1071].
In the Middle East, Iran has received its first doses of Sputnik V and the United Arab Emirates is conducting phase III trials [1071].
In Asia, while China’s vaccine candidates are favored, the Philippines, Nepal, and Uzbekistan have sought Sputnik V doses [1072,1072].
In total, the RDIF claims to have received orders totalling 1.2 billion doses by over 50 countries worldwide [1072] and at least 18 countries are currently administering Sputnik V around the globe [1031].
Sputnik V has been an attractive vaccine for many countries due to its relatively low price, high efficacy, and its favorable storage conditions.
For some countries, Russia and China have also been more palatable politically than vaccine suppliers in the West [1071,1073].
For others, the delays in the distribution of the other, more-favored candidates has been a motivating factor for pursuing the Sputnik V and Chinese alternatives [1073; https://www.euronews.com/2021/02/24/san-marino-buys-russia-s-sputnik-v-after-eu-vaccine-delivery-delays].
Additionally, Germany has stated that if Sputnik V were approved by EMA, it would be considered by the E.U. [1074].
Russia is developing other vaccine candidates and has approved a third vaccine, CoviVac, which is an inactivated vaccine produced by the Chumakov Centre in Moscow, despite the fact the clinical trials have yet to begin [1075].
In Asia, China and India are the main COVID-19 vaccination developers and providers.
In India, the Covaxin vaccine produced by Bharat Biotech received emergency authorization on January 3rd, 2021, despite the lack of phase III data until March 3rd [1076,1077].
Following the release of the phase III data indicating 81% efficacy, Zimbabwe authorized the use of Covaxin [1078].
In February, 2021, Bharat Biotech received approval from Indian officials to commence a phase I study of an intranasal chimpanzee-adenovirus (ChAd) vectored SARS-CoV-2-S vaccine called BBV154 [1079].
Notably, Novavax has signed an agreement with the Serum Institute of India allowing them to produce up to 2 billion doses a year [1080].
Novavax has also signed agreements with the U.K., Canada, Australia, and South Korea [1081] and has projected that they will supply 1.1 billion doses to COVAX who will distribute the vaccines to countries with disadvantaged access to vaccine supplies [1053].
India has vaccinated approximately 24 million people [1032].
This has been achieved by mainly using the AstraZeneca-University of Oxford vaccine, known as Covishield in India, which is also produced by the Serum Institute of India, and using India’s own Covaxin vaccine [1082].
India has also shipped approximately 58 million COVID-19 vaccines to 66 countries [1083]
Considering India produces approximately 60% of the world’s vaccines prior to the pandemic, it is no surprise that several other vaccine candidates are under development.
These include ZyCov-Di a DNA vaccine produced by Zydus Cadila, HGCO19 India’s first mRNA vaccine produced by Genova and HDT Biotech Corporation (USA), and the Bio E subunit vaccine produced by Biological E in collaboration with US-based Dynavax and the Baylor College of Medicine [1082].
In China, the Sinopharm-Beijing Institute vaccine, the Sinopharm-Wuhan Institute of Biological Products vaccine, the Sinovac Biotech (CoronaVac) vaccine, and CanSino Biologics vaccine are the main vaccines being distributed.
The Sinopharm-Beijing vaccine has been distributed to at least 16 countries.
This vaccine is currently approved for use in Bahrain, China, and the United Arab Emirates, but has been granted emergency use in Argentina, Cambodia, Egypt, Guyana, Hungary, Iran, Iraq, Jordan, Nepal, Pakistan, Peru, Venezuela, and Zimbabwe, with limited use in both Serbia and the Seychelles [1084].
The Sinovac vaccine, CoronaVac, has been approved for use in China, and has been granted emergency use in Azerbaijan, Brazil, Cambodia, Chile, Colombia, Ecuador, Hong Kong, Indonesia, Laos, Malaysia, Mexico, Philippines, Thailand, Turkey, Ukraine, and Uruguay [1085].
Sinovac has reported that their platform now has the capacity to provide up to a billion doses [1085].
Indeed, Sinovac and Sinopharm have estimated that they will be able to produce 2 billion doses by the end of 2021, and they have been able to distribute vaccines as aid to the Philippines and Pakistan [1086].
In contrast, the Sinopharm-Wuhan vaccine, which has been approved for use in China since February 25th, 2021, has been distributed almost exclusively within China, with limited supplies distributed to the United Arab Emirates [1087].
On the same date, the CanSino vaccine was approved for use in China and has been granted emergency use in Mexico and Pakistan, which were two participating countries in the CanSino phase III trials [1088].
However, the vaccine approval and distribution processes in China have come under increased scrutiny from other nations.
China was criticized for administering vaccines to thousands of government officials and state-owned businesses in September 2020, prior to the completion of phase III clinical trials [1052].
The behavior of Chinese officials has also come into question due to misinformation campaigns questioning the safety of Western vaccine candidates such as Moderna and Pfizer-BioNTech in a way that is intended to highlight the benefits of their own vaccine candidates [1086].
Furthermore, delays in vaccine distribution have also caused issues, particularly in Turkey where 10 million doses of Sinovac were due to arrive by December 2020, but instead only 3 million were delivered in early January [1086].
Similar delays and shortages of doses promised have been reported by officials in the Philippines, Egypt, Morocco, and the United Arab Emirates [1089,1090].
This will be concerning to China who have vaccine contracts for millions of doses with Indonesia (>100 million), Brazil (100 million), Chile (60 million), Turkey (50 million), Egypt (40 million) and many others [1090].
Globally, North America currently leads the world vaccination rates (13.8 per 100 people) followed by Europe (8.2 per 100), South America (3.1 per 100), Asia (1.9 per 100), Africa (0.3 per 100), and Oceania (0.1 per 100) are trailing behind [1031].
Considering the wealthy nations of North America and Europe have secured most of the limited COVID-19 vaccine stocks [1091], it is likely that low- and middle-income countries will face further competition with Western countries for vaccine availability.
While South Africa and Zimbabwe have their own vaccination programs, many other African nations will be reliant on the COVID-19 Vaccines Global Access (COVAX) Facility, who have promised 600 million doses to the continent [1092].
COVAX is a multilateral initiative as part of the Access to COVID-19 Tools (ACT) Accelerator coordinated by the WHO, Gavi The Vaccine Alliance, and the Coalition for Epidemic Preparedness Innovations (CEPI), the latter two of which are supported by the Bill and Melinda Gates Foundation.
Their intention is to accelerate the development of COVID-19 vaccines, diagnostics, and therapeutics and to ensure the equitable distribution of vaccines to low- and middle-income countries [1093,1094].
COVAX invested in several vaccine programs to ensure they would have access to successful vaccine candidates [1095].
The COVAX plan ensured that all participating countries would be allocated vaccines in proportion to their population sizes.
Once each country has received vaccine doses to account for 20% of their population, the country’s risk profile will determine its place in subsequent phases of vaccine distribution.
However, several limitations of this framework exist, including that the COVAX scheme seems to go against the WHO’s own ethical principles of human well-being, equal respect, and global equity, and that other frameworks might have been more suitable, as is discussed elsewhere [1096].
Furthermore, COVAX is supposed to allow poorer countries access to affordable vaccines, but the vaccines are driven by publicly traded companies that are required to make a profit [1033].
In any case, COVAX provides access to COVID-19 vaccines that may otherwise have been difficult for some countries to obtain.
COVAX aims to distribute 2 billion vaccine doses globally by the end of 2021 [1097].
COVAX may also receive additional donations of doses from Western nations who purchased surplus vaccines in the race to vaccinate their populations, which will be a welcome boost to the vaccination programs of low- and middle-income countries [1098].
As of March, 2021, 9 African countries have received vaccines and at least 11 other nations have begun vaccinations via COVAX, aid from other countries, or their own agreements with producers [1092,1099].
However, much further progress is required when only 0.3 per 100 people have been vaccinated in Africa [1031].
7.13 Discussion
Additionally, major advances in vaccines using mRNA and adenoviruses that have led to three vaccines becoming available or close to becoming available in late 2020 (Figure 4).
Though some concerns remain about the duration of sustained immunity for convalescents, vaccine development efforts are ongoing and show initial promising results.
The Moderna trial, for example, reported that the neutralizing activity in participants who received two doses of the vaccine was similar to that observed in convalescent plasma.
One of the two mRNA vaccines, Pfizer and BioNTech’s BNT162b2, has been issued an EUA for patients as young as 16 [1100], while ModernaTX has begun a clinical trial to assess its mRNA vaccine in adolescents ages 12 to 18 [1101].
8 Social Factors Influencing COVID-19 Exposure and Outcomes
8.1 Social Factors Influencing COVID-19 Outcomes
In addition to understanding the fundamental biology of the SARS-CoV-2 virus and COVID-19, it is critical to consider how the broader environment can influence both COVID-19 outcomes and efforts to develop and implement treatments for the disease.
The evidence clearly indicates that social environmental factors are critical determinants of individuals’ and communities’ risks related to COVID-19.
There are distinct components to COVID-19 susceptibility, and an individual’s risk can be elevated at one or all stages from exposure to recovery/mortality: an individual may be more likely to be exposed to the virus, more likely to get infected once exposed, more likely to have serious complications once infected, and be less likely to receive adequate care once they are seriously ill.
The fact that differences in survival between Black and white patients were no longer significant after controlling for comorbidities and socioeconomic status (type of insurance, neighborhood deprivation score, and hospital where treatment was received) in addition to sex and age [1102] underscores the relevance of social factors to understanding mortality differences between racial and ethnic groups.
Moreover, the Black patients were younger and more likely to be female than white patients, yet still had a higher mortality rate without correction for the other variables [1102].
Here, we outline a few systemic reasons that may exacerbate the COVID-19 pandemic in communities of color.
8.2 Factors Observed to be Associated with Susceptibility
As COVID-19 has spread into communities around the globe, it has become clear that the risks associated with this disease are not equally shared by all individuals or all communities.
Significant disparities in outcomes have led to interest in the demographic, biomedical, and social factors that influence COVID-19 severity.
Untangling the factors influencing COVID-19 susceptibility is a complex undertaking.
Among patients who are admitted to the hospital, outcomes have generally been poor, with rates of admission to the intensive care unit (ICU) upwards of 15% in both Wuhan, China and Italy [37,1103,1104].
However, hospitalization rates vary by location [1105].
This variation may be influenced by demographic (e.g., average age in the area), medical (e.g., the prevalence of comorbid conditions such as diabetes), and social (e.g., income or healthcare availability) factors that vary geographically.
Additionally, some of the same factors may influence an individual’s probability of exposure to SARS-CoV-2, their risk of developing a more serious case of COVID-19 that would require hospitalization, and their access to medical support.
As a result, quantifying or comparing susceptibility among individuals, communities, or other groups requires consideration of a number of complex phenomena that intersect across many disciplines of research.
In this section, the term “risk factor” is used to refer to variables that are statistically associated with more severe COVID-19 outcomes.
Some are intrinsic characteristics that have been observed to carry an association with variation in outcomes, whereas others may be more functionally linked to the pathophysiology of COVID-19.
8.2.1 Patient Traits Associated with Increased Risk
Two traits that have been consistently associated with more severe COVID-19 outcomes are male sex and advanced age (typically defined as 60 or older, with the greatest risk among those 85 and older [1106]).
In the United States, males and older individuals diagnosed with COVID-19 were found to be more likely to require hospitalization [1107,1108].
A retrospective study of hospitalized Chinese patients [38] found that a higher probability of mortality was associated with older age, and world-wide, population age structure has been found to be an important variable for explaining differences in outbreak severity [1109].
The CFR for adults over 80 has been estimated upwards of 14% or even 20% [1110].
Male sex has also been identified as a risk factor for severe COVID-19 outcomes, including death [1111,1112,1113].
Early reports from China and Europe indicated that even though the case rates were similar across males and females, males were at elevated risk for hospital admission, ICU admission, and death [1112], although data from some US states indicates more cases among females, potentially due to gender representation in care-taking professions [1114].
In older age groups (e.g., age 60 and older), comparable absolute numbers of male and female cases actually suggests a higher rate of occurrence in males, due to increased skew in the sex ratio [1112].
Current estimates based on worldwide data suggest that, compared to females, males may be 30% more likely to be hospitalized, 80% more likely to be admitted to the ICU, and 40% more likely to die as a result of COVID-19 [1113].
There also may be a compounding effect of advanced age and male sex, with differences time to recovery worst for males over 60 years old relative to female members of their age cohort [1115].
Both of these risk factors can be approached through the lens of biology.
The biological basis for greater susceptibility with age is likely linked to the prevalence of extenuating health conditions such as heart failure or diabetes [1110].
Several hypotheses have been proposed to account for differences in severity between males and females.
For example, some evidence suggests that female sex hormones may be protective [1112,1114].
ACE2 expression in the kidneys of male mice was observed to be twice as high as that of females, and a regulatory effect of estradiol on ACE2 expression was demonstrated by removing the gonads and then supplementing with estradiol [1114,1116].
Other work in mice has shown an inverse association between mortality due to SARS-CoV-1 and estradiol, suggesting a protective role for the sex hormone [1114].
Similarly, evidence suggests that similar patterns might be found in other tissues.
A preliminary analysis identified higher levels of ACE2 expression in the myocardium of male patients with aortic valve stenosis showed than female patients, although this pattern was not found in controls [1112].
Additionally, research has indicated that females respond to lower doses than males of heart medications that act on the Renin angiotensin aldosterone system (RAAS) pathway, which is shared with ACE2 [1112].
Additionally, several components of the immune response, including the inflammatory response, may differ in intensity and timing between males and females [1114,1116].
This hypothesis is supported by some preliminary evidence showing that female patients who recovered from severe COVID-19 had higher antibody titers than males [1114].
Sex steroids can also bind to immune cell receptors to influence cytokine production [1112].
Additionally, social factors may influence risks related to both age and sex: for example, older adults are more likely to live in care facilities, which have been a source for a large number of outbreaks [1117], and gender roles may also influence exposure and/or susceptibility due to differences in care-taking and/or risky behaviors (e.g., caring for elder relatives and smoking, respectively) [1112] among men and women (however, it should be noted that both transgender men and women are suspected to be at heightened risk [1118].)
8.2.2 Comorbid Health Conditions
A number of pre-existing or comorbid conditions have repeatedly been identified as risk factors for more severe COVID-19 outcomes.
Several underlying health conditions were identified at high prevalence among hospitalized patients, including obesity, diabetes, hypertension, lung disease, and cardiovascular disease [1105].
Higher Sequential Organ Failure Assessment (SOFA) scores have been associated with a higher probability of mortality [38], and comorbid conditions such as cardiovascular and lung disease as well as obesity were also associated with an increased risk of hospitalization and death, even when correcting for age and sex [1111].
Diabetes may increase the risk of lengthy hospitalization [1119] or of death [1119,1120].
[1121] and [1122] discuss possible ways in which COVID-19 and diabetes may interact.
Obesity also appears to be associated with higher risk of severe outcomes from SARS-CoV-2 [1123,1124].
Obesity is considered an underlying risk factor for other health problems, and the mechanism for its contributions to COVID-19 hospitalization or mortality is not yet clear [1125].
Dementia and cancer were also associated with the risk of death in an analysis of a large number (more than 20,000) COVID-19 patients in the United Kingdom [1111].
It should be noted that comorbid conditions are inextricably tied to age, as conditions tend to be accumulated over time, but that the prevalence of individual comorbidities or of population health overall can vary regionally [1126].
Several comorbidities that are highly prevalent in older adults, such as COPD, hypertension, cardiovascular disease, and diabetes, have been associated with CFRs upwards of 8% compared to an estimate of 1.4% in people without comorbidities [1110,1127].
Therefore, both age and health are important considerations when predicting the impact of COVID-19 on a population [1126].
However, other associations may exist, such as patients with sepsis having higher SOFA scores – in fact, SOFA was developed for the assessment of organ failure in the context of sepsis, and the acronym originally stood for Sepsis-Related Organ Failure Assessment [1128,1129].
Additionally, certain conditions are likely to be more prevalent under or exacerbated by social conditions, especially poverty, as is discussed further below.
8.2.3 Ancestry
A number of studies have suggested associations between individuals’ racial and ethnic backgrounds and their COVID-19 risk.
In particular, Black Americans are consistently identified as carrying a higher burden of COVID-19 than white Americans [1107,1108], with differences in the rates of kidney complications from COVID-19 particularly pronounced [110].
Statistics from a number of cities indicate significant discrepancies between the proportion of COVID-19 cases and deaths in Black Americans relative to their representation in the general population [1130].
In addition to Black Americans, disproportionate harm and mortality from COVID-19 has also been noted in Latino/Hispanic Americans and in Native American and Alaskan Native communities, including the Navajo nation [1131,1132,1133,1134,1135,1136].
In Brazil, indigenous communities likewise carry an increased burden of COVID-19 [1137].
In the United Kingdom, nonwhite ethnicity (principally Black or South Asian) was one of several factors found to be associated with a higher risk of death from COVID-19 [1138].
From a genetic standpoint, it is highly unlikely that ancestry itself predisposes individuals to contracting COVID-19 or to experiencing severe COVID-19 outcomes.
Examining human genetic diversity indicates variation over a geographic continuum, and that most human genetic variation is associated with the African continent [1139].
African-Americans are also a more genetically diverse group relative to European-Americans, with a large number of rare alleles and a much smaller fraction of common alleles identified in African-Americans [1140].
Therefore, the idea that African ancestry (at the continent level) might convey some sort of genetic risk for severe COVID-19 contrasts with what is known about worldwide human genetic diversity [1141].
The possibility for genetic variants that confer some risk or some protection remains possible, but has not been widely explored, especially at a global level.
Research in Beijing of a small number (n=80) hospitalized COVID-19 patients revealed an association between severe COVID-19 outcomes and homozygosity for an allele in the interferon-induced transmembrane protein 3 (IFITM3) gene, which was selected as a candidate because it was previously found to be associated with influenza outcomes in Chinese patients [1142].
Genetic factors may also play a role in the risk of respiratory failure for COVID-19 [1143,1144,1145].
However, genetic variants associated with outcomes within ancestral groups are far less surprising than genetic variants explaining outcomes between groups.
Alleles in ACE2 and TMPRSS2 have been identified that vary in frequency among ancestral groups [1146], but whether these variants are associated with COVID-19 susceptibility has not been explored.
Instead, examining patterns of COVID-19 susceptibility on a global scale that suggest that social factors are of primary importance in predicting mortality.
Reports from several sub-Saharan African countries have indicated that the effects of the COVID-19 pandemic have been less severe than expected based on the outbreaks in China and Italy.
In Kenya, for example, estimates of national prevalence based on testing blood donors for SARS-CoV-2 antibodies were consistent with 5% of Kenyan adults having recovered from COVID-19 [1147].
This high seroprevalence of antibodies lies in sharp contrast to the low number of COVID-19 fatalities in Kenya, which at the time was 71 out of 2093 known cases [1147].
Likewise, a serosurvey of health care workers in Blantyre City, Malawi reported an adjusted antibody prevalence of 12.3%, suggesting that the virus had been circulating more widely than thought and that the death rate was up eight times lower than models had predicted [1148].
While several possible hypotheses for the apparent reduced impact of COVID-19 on the African continent are being explored, such as young demographics in many places [1149], these reports present a stark contrast to the severity of COVID-19 in Americans and Europeans of African descent.
Additionally, ethnic minorities in the United Kingdom also tend to be younger than white British living in the same areas, yet the burden of COVID-19 is still more serious for minorities, especially people of Black Caribbean ancestry, both in absolute numbers and when controlling for age and location [1150].
Furthermore, the groups in the United States and United Kingdom that have been identified as carrying elevated COVID-19 burden, namely Black American, indigenous American, and Black and South Asian British, are quite distinct in their position on the human ancestral tree.
What is shared across these groups is instead a history of disenfranchisement under colonialism and ongoing systematic racism.
A large analysis of over 11,000 COVID-19 patients hospitalized in 92 hospitals across U.S. states revealed that Black patients were younger, more often female, more likely to be on Medicaid, more likely to have comorbidities, and came from neighborhoods identified as more economically deprived than white patients [1102].
This study reported that when these factors were accounted for, the differences in mortality between Black and white patients were no longer significant.
Thus, the current evidence suggests that the apparent correlations between ancestry and health outcomes must be examined in the appropriate social context.
8.3 Environmental Influences on Susceptibility
8.3.1 Exposure to COVID-19
Social distancing has emerged as one of the main social policies used to manage the COVID-19 epidemic in many countries.
Many governments issued stay-at-home orders, especially in the initial months of the crisis.
However, data clearly indicates that these orders impacted different socioeconomic groups differently.
In U.S. counties with and without stay-at-home orders, smartphone tracking indicated a significant decrease in the general population’s mobility in April relative to February through March of 2020 (-52.3% and -60.8%, respectively) [1151].
A linear relationship was observed between counties’ reduction in mobility and their wealth and health, as measured by access to health care, food security, income, space, and other factors [1151].
Counties with greater reductions in mobility were also found to have much lower child poverty and household crowding and to be more racially segregated, and to have fewer youth and more elderly residents [1151].
Similar associations between wealth and decreased mobility were observed in cellphone GPS data from Colombia, Indonesia, and Mexico collected between January and May 2020 [1152], as well as in a very large data set from several US cities [1153].
These disparities in mobility are likely to be related to the role that essential workers have played during the pandemic.
Essential workers are disproportionately likely to be female, people of color, immigrants, and to have an income below 200% of the poverty line [1154].
Black Americans in particular are over-represented among front-line workers and in professions where social distancing is infeasible [1155].
Health care work in particular presents an increased risk of exposure to SARS-CoV-2 [1155,1156,1157,1158,1159].
In the United Kingdom, (South) Asians are more likely than their white counterparts to be medical professionals [1150], although BAME medical professionals are still disproportionately represented in the proportion of National Health Service staff deaths [1160].
Similar trends have been reported for nurses, especially nurses of color, in the United States [1161].
Furthermore, beyond the risks associated with work itself, use of public transportation may also impact COVID-19 risk [1162].
The socioeconomic and racial/ethnic gaps in who is working on the front lines of the pandemic make it clear that socioeconomic privilege is likely to decrease the probability of exposure to SARS-CoV-2.
Increased risk of exposure can also arise outside the workplace.
Nursing homes and skilled nursing facilities received attention early on as high-risk locations for COVID-19 outbreaks [1163].
Prisons and detention centers also confer a high risk of exposure or infection [1164,1165].
Populations in care facilities are largely older adults, and in the United States, incarcerated people are more likely to be male and persons of color, especially Black [1166].
Additionally, multi-generational households are less common among non-Hispanic white Americans than people of other racial and ethnic backgrounds [1167], increasing the risk of exposure for more susceptible family members.
Analysis suggests that household crowding may also be associated with increased risk of COVID-19 exposure [1151], and household crowding is associated with poverty [1168].
Forms of economic insecurity like housing insecurity, which is associated with poverty and more pronounced in communities subjected to racism [1169,1170], would be likely to increase household crowding and other possible sources of exposure.
As a result, facets of systemic inequality such as mass incarceration of Black Americans and poverty are likely to increase the risk of exposure outside of the workplace.
8.3.2 Severity of COVID-19 Following Exposure
Following exposure to SARS-CoV-2, the likelihood that an individual develops COVID-19 and the severity of the disease presentation can be influenced by a number of social factors.
As discussed above, a number of patient characteristics are associated with the likelihood of severe COVID-19 symptoms.
In some cases, these trends run counter to those expected given rates of exposure: for example, although women are more likely to be exposed, men are more likely to be diagnosed with, hospitalized from, or die from COVID-19 [1114].
In the case of comorbid conditions and racial/ethnic demographics, however, social factors are highly likely to modulate or at least influence the apparent association between these traits and the increased risk from COVID-19.
In particular, the comorbidities and racial/ethnic correlates of severe COVID-19 outcomes suggest that poverty confers additional risk for COVID-19.
In order to explore the relationship between poverty and COVID-19 outcomes, it is necessary to consider how poverty impacts biology.
In particular, we focus on the United States and the United Kingdom.
Comorbidities that increase risk for COVID-19, including obesity, type II diabetes, hypertension, and cardiovascular disease, are known to be intercorrelated [1171].
Metabolic conditions related to heightened inflammation, like obesity, type II diabetes, and hypertension, are more strongly associated with negative COVID-19 outcomes than other comorbid conditions, such as chronic heart disease [1172].
As discussed above, dysregulated inflammation characteristic of cytokine release syndrome is one of the greatest concerns for COVID-19-related death.
Therefore, it is possible that chronic inflammation characteristic of these metabolic conditions predisposes patients to COVID-19-related death [1172].
The association between these diseases and severe COVID-19 outcomes is a concern from a health equity perspective because poverty exposes people to “obesogenic” conditions [1173] and is therefore unsurprisingly associated with higher incidence of obesity and associated disorders [1174].
Furthermore, cell phone GPS data suggests that lower socioeconomic status may also be associated with decreased access to healthy food choices during the COVID-19 pandemic [1175,1176], suggesting that health-related risk factors for COVID-19 may be exacerbated as the pandemic continues [1177].
Chronic inflammation is a known outcome of chronic stress (e.g., [1178,1179,1180,1181]).
Therefore, the chronic stress of poverty is likely to influence health broadly (as summarized in [1182]) and especially during the stress of the ongoing pandemic.
A preprint [1183] provided observational evidence that geographical areas in the United States that suffer from worse air pollution by fine particulate matter have also suffered more COVID-19 deaths per capita, after adjusting for demographic covariates.
Although lack of individual-level exposure data and the impossibility of randomization make it difficult to elucidate the exact causal mechanism, this finding would be consistent with similar findings for all-cause mortality (e.g., [1184]).
Exposure to air pollution is associated with both poverty (e.g., [1185]) and chronic inflammation [1186].
Other outcomes of environmental racism, such as the proximity of abandoned uranium mines to Navajo land, can also cause respiratory illnesses and other health issues [1136].
Similarly, preliminary findings indicate that nutritional status (e.g., vitamin D deficiency [779]) may be associated with COVID-19 outcomes, and reduced access to grocery stores and fresh food often co-occurs with environmental racism [1136,1187].
Taken together, the evidence suggests that low-income workers who face greater exposure to SARS-CoV-2 due to their home or work conditions are also more likely to face environmental and social stressors associated with increased inflammation, and therefore with increased risk from COVID-19.
In particular, structural racism can play an important role on disease severity after SARS-CoV-2 exposure, due to consequences of racism which include an increased likelihood of poverty and its associated food and housing instability.
COVID-19 can thus be considered a “syndemic”, or a synergistic interaction between several epidemics [1188].
As a result, it is not surprising that people from minoritized backgrounds and/or with certain pre-existing conditions are more likely to suffer severe effects of COVID-19, but these “risk factors” are likely to be causally linked to poverty [1189].
8.3.3 Access to Treatment
Finally, COVID-19 outcomes can be influenced by access to healthcare.
Receiving care for COVID-19 can, but does not always, include receiving a positive test for the SARS-CoV-2 virus.
For example, it is common to see treatment guidelines for suspected cases regardless of whether the presence of SARS-CoV-2 has been confirmed (e.g., [1190]).
Whether and where a patient is diagnosed can depend on their access to testing, which can vary both between and within countries.
In the United States, it is not always clear whether an individual will have access to free testing [1191,1192].
The concern has been raised that more economic privilege is likely to correspond to increased access to testing, at least within the United States [1193].
This is supported by the fact that African Americans seem to be more likely to be diagnosed in the hospital, while individuals from other groups were more likely to have been diagnosed in ambulatory settings in the community [1107].
Any delays in treatment are a cause for concern [1193], which could potentially be increased by an inability to acquire testing because in the United States, insurance coverage for care received can depend on a positive test [1194].
Another important question is whether patients with moderate to severe cases are able to access hospital facilities and treatments, to the extent that they have been identified.
Early findings from China as of February 2020 suggested the COVID-19 mortality rate to be much lower in the most developed regions of the country [1195], although reported mortality is generally an estimate of CFR, which is dependent on rates of testing.
Efforts to make treatment accessible for all confirmed and suspected cases of COVID-19 in China are credited with expanding care to people with fewer economic resources [1196].
In the United States, access to healthcare varies widely, with certain sectors of the workforce less likely to have health insurance; many essential workers in transportation, food service, and other frontline fields are among those likely to be uninsured or underinsured [1193].
As of 2018, Hispanic Americans of all races were much less likely to have health insurance than people from non-Hispanic backgrounds [1197].
Therefore, access to diagnostics and care prior to the development of severe COVID-19 is likely to vary depending on socioeconomic and social factors, many of which overlap with the risks of exposure and of developing more severe COVID-19 symptoms.
This discrepancy ties into concerns about broad infrastructural challenges imposed by COVID-19.
A major concern in many countries has been the saturation of healthcare systems due to the volume of COVID-19 hospitalizations (e.g., [260]).
Similarly, there have been shortages of supplies such as ventilators that are critical to the survival of many COVID-19 patients, leading to extensive ethical discussions about how to allocate limited resources among patients [1198,1199,1200,1201].
Although it is generally considered unethical to consider demographic factors such as age, sex, race, or ethnicity while making such decisions, and ideally this information would not be shared with triage teams tasked with allocating limited resources among patients [1202], there are substantial concerns about implicit and explicit biases against older adults [1203], premature infants [1204], and people with disabilities or comorbidities [1202,1205,1206].
Because of the greater burden of chronic disease in populations subjected to systemic racism, algorithms intended to be blind to race and ethnicity could, in fact, reinforce systemic inequalities caused by structural racism [1207,1208,1209].
Because of this inequality, it has been argued that groups facing health disparities should be prioritized by these algorithms [1210].
This approach would carry its own ethical concerns, including the fact that many resources that need to be distributed do not have well-established risks and benefits [1210].
As the pandemic has progressed, it has become clear that ICU beds and ventilators are not the only limited resources that needs to be allocated, and, in fact, the survival rate for patients who receive mechanical ventilation is lower than these discussions would suggest [1211].
Allocation of interventions that may reduce suffering, including palliative care, has become critically important [1211,1212].
The ambiguities surrounding the risks and benefits associated with therapeutics that have been approved under emergency use authorizations also present ethical concerns related to the distribution of resources [1210].
For example, remdesivir, discussed above, is currently available for the treatment of COVID-19 under compassionate use guidelines and through expanded access programs, and in many cases has been donated to hospitals by Gilead [1213,1214].
Regulations guiding the distribution of drugs in situations like these typically do not address how to determine which patients receive them [1214].
Prioritizing marginalized groups for treatment with a drug like remdesivir would also be unethical because it would entail disproportionately exposing these groups to a therapeutic that may or not be beneficial [1210].
On the other hand, given that the drug is one of the most promising treatments available for many patients, using a framework that tacitly feeds into structural biases would also be unethical.
At present, the report prepared for the Director of the CDC by Ethics Subcommittee of the CDC fails to address the complexity of this ethical question given the state of structural racism in the United States, instead stating that “prioritizing individuals according to their chances for short-term survival also avoids ethically irrelevant considerations, such as race or socioeconomic status” [1215].
In many cases, experimental therapeutics are made available only through participation in clinical trials [1216].
However, given the history of medical trials abusing minority communities, especially Black Americans, there is a history of unequal representation in clinical trial enrollment [1216].
As a result, the standard practice of requiring enrollment in a clinical trial in order to receive experimental treatment may also reinforce patterns established by systemic racism.
8.3.4 Access to and Representation in Clinical Trials
Experimental treatments are often made available to patients primarily or even exclusively through clinical trials.
The advantage of this approach is that clinical trials are designed to collect rigorous data about the effects of a treatment on patients.
The disadvantage is that access to clinical trials is not equal among all people who suffer from a disease.
Two important considerations that can impact an individual’s access to clinical trials are geography and social perceptions of clinical trials.
For the first, the geographic distribution of trial recruitment efforts are typically bounded and can vary widely among difference locations, and for the second, the social context of medical interactions can impact strategies for and the success of outreach to different communities.
Differential access to clinical trials raises concerns because it introduces biases that can influence scientific and medical research on therapeutics and prophylactics broadly.
Concerns about bias in clinical trials need to address both trial recruitment and operation.
In the present crisis, such biases are particularly salient because COVID-19 is a disease of global concern.
Treatment is needed by people all over the world, and clinical research that characterizes treatment outcomes in a variety of populations is critically important.
Global representation in clinical trials is important to ensuring that experimental treatments are available equally to COVID-19 patients who may need them.
The advantage to a patient of participation in a clinical trial is that they may receive an experimental treatment they would not have been able to access otherwise.
The potential downsides of participation include that the efficacy and side effects of such treatments are often poorly characterized and that patients who enroll in clinical trials will in some cases run the risk of being assigned to a placebo condition where they do not receive the treatment but miss out on opportunities to receive other treatments.
The benefits and burdens of clinical trials therefore need to be weighed carefully to ensure that they don’t reinforce existing health disparities.
The WHO Director‐General Tedros Adhanom Ghebreyesus stated his condemnation of utilizing low and middle income countries as test subjects for clinical trials, yet having highly developed countries as the majority of clinical trial representation is also not the answer [1217].
Figure 7 showcases two choropleths detailing COVID-19 clinical trial recruitment by country.
China, the United States, and France are among the countries with the most clinical trial recruiting for trials with single-country enrollment.
Many countries have little to no clinical trial recruiting, with the continents of Africa and South America much less represented than Asia, Europe, and North America.
Trials that recruit across multiple countries do appear to broaden geographic representation, but these trials seem to be heavily dominated by the United States and European Union.
Figure 7:Geographic distribution of COVID-19 clinical trials.
The density of clinical trials is reported at the country level.
As of November 9, 2020, there are 6,417 trials in the University of Oxford Evidence-Based Medicine Data Lab’s COVID-19 TrialsTracker [405], of which 3,706 are interventional.
The top figure demonstrates the density of interventional trials recruiting only from a singular country, while the bottom shows the distribution of recruitment for interventional trials that involve more than one country.
A few different concerns arise from this skewed geographic representation in clinical trial recruitment.
First, treatments such as remdesivir that are promising but primarily available to clinical trial participants are unlikely to be accessible by people in many countries.
Second, it raises the concern that the findings of clinical trials will be based on participants from many of the wealthiest countries, which may lead to ambiguity in whether the findings can be extrapolated to COVID-19 patients elsewhere.
Especially with the global nature of COVID-19, equitable access to therapeutics and vaccines has been a concern at the forefront of many discussions about policy (e.g., [1218], yet data like that shown in Figure 7 demonstrates that accessibility is likely to be a significant issue.
Another concern with the heterogeneous international distribution of clinical trials is that the governments of countries leading these clinical trials might prioritize their own populations once vaccines are developed, causing unequal health outcomes [1219].
Additionally, even within a single state in the United States (Maryland), geography was found to influence the likelihood of being recruited into or enrolled in a clinical trial, with patients in under-served rural areas less likely to enroll [1220].
Thus, geography both on the global and local levels may influence when treatments and vaccines are available and who is able to access them.
Efforts such as the African Union’s efforts to coordinate and promote vaccine development [1221] are therefore critical to promoting equity in the COVID-19 response.
Even when patients are located within the geographic recruitment area of clinical trials, however, there can still be demographic inequalities in enrollment.
When efforts are made to ensure equal opportunity to participate in clinical trials, there is no significant difference in participation among racial/ethnic groups [1222].
However, within the United States, real clinical trial recruitment numbers have indicated for many years that racial minorities, especially African-Americans, tend to be under-represented (e.g., [1223,1224,1225,1226]).
This trend is especially concerning given the disproportionate impact of COVID-19 on African-Americans.
Early evidence suggests that the proportion of Black, Latinx, and Native American participants in clinical trials for drugs such as remdesivir is much lower than the representation of these groups among COVID-19 patients [1227].
One proposed explanation for differences among racial and ethnic groups in clinical trial enrollment refers to different experiences in healthcare settings.
While some plausible reasons for the disparity in communication between physicians and patients could be a lack of awareness and education, mistrust in healthcare professionals, and a lack of health insurance [1222], a major concern is that patients from certain racial and ethnic groups are marginalized even while seeking healthcare.
In the United States, many patients experience “othering” from physicians and other medical professionals due to their race or other external characteristics such as gender (e.g., [1228]).
Many studies have sought to characterize implicit biases in healthcare providers and whether they affect their perceptions or treatment of patients.
A systematic review that examined 37 such studies reported that most (31) identified racial and/or ethnic biases in healthcare providers in many different roles, although the evidence about whether these biases translated to different attitudes towards patients was mixed [1229], with similar findings reported by a second systematic review [1230].
However, data about real-world patient outcomes are very limited, with most studies relying on clinical vignette-based exercises [1229], and other analyses suggest that physician implicit bias could impact the patient’s perception of the negativity/positivity of the interaction regardless of the physician’s explicit behavior towards the patient [1231].
Because racism is a common factor in both, negative patient experiences with medical professionals are likely to compound other issues of systemic inequality, such as a lack of access to adequate care, a lack of insurance, or increased exposure to SARS-CoV-2 [1232].
Furthermore, the experience of being othered is not only expected to impact patients’ trust in and comfort with their provider, but also may directly impact whether or not the patient is offered the opportunity to participate in a clinical trial at all.
Some studies suggest communication between physicians and patients impacts whether or not a physician offers a patient participation in a clinical trial.
For example, researchers utilized a linguistic analysis to assess mean word count of phrases related to clinical trial enrollment, such as voluntary participation, clinical trial, etc. [1222].
The data indicated that the mean word count of the entire visit was 1.5 times more for white patients in comparison to Black patients.
In addition, the greatest disparity between white and Black patients’ experience was the discussion of risks, with over 2 times as many risk-related words spoken with white patients than Black patients [1222].
The trends observed for other clinical trials raise the concern that COVID-19 clinical trial information may not be discussed as thoroughly or as often with Black patients compared to white patients.
These discrepancies are especially concerning given that many COVID-19 treatments are being or are considered being made available to patients prior to FDA approval through Emergency Use Authorizations.
In the past, African-Americans have been over-represented relative to national demographics in use of the FDA’s Exception From Informed Consent (EFIC) pathway [1233].
Through this pathway, people who are incapacitated can receive an experimental treatment even if they are not able to consent and there is not sufficient time to seek approval from an authorized representative.
This pathway presents concerns, however, when it is considered in the context of a long history of systematic abuses in medical experimentation where informed consent was not obtained from people of color, such as the Tuskegee syphilis experiments [1234].
While the goal of EFIC approval is to provide treatment to patients who urgently need it, the combination of the ongoing legacy of racism in medicine renders this trend concerning.
With COVID-19, efforts to prioritize people who suffer from systemic racism are often designed with the goal of righting some of these inequalities (e.g., [1235]), but particular attention to informed consent will be imperative in ensuring these trials are ethical given that the benefits and risks of emerging treatments are still poorly characterized.
Making a substantial effort to run inclusive clinical trials is also important because of the possibility that racism could impact how a patient responds to a treatment.
For example, as discussed above, dexamethasone has been identified as a promising treatment for patients experiencing cytokine release syndrome, but the mechanism of action is tied to the stress response.
A study from 2005 reported that Black asthma patients showed reduced responsiveness to dexamethasone in comparison to white patients and suggested Black patients might therefore require higher doses of the drug [1236].
In the context of chronic stress caused by systemic racism, this result is not surprising: chronic stress is associated with dysregulated production of glucocorticoids [1237] and glucocorticoid receptor resistance [1238].
However, it underscores the critical need for treatment guidelines to take into account differences in life experience, which would be facilitated by the recruitment of patients from a wide range of backgrounds.
Attention to the social aspects of clinical trial enrollment must therefore be an essential component of the medical research community’s response to COVID-19.
8.4 Conclusions and Future Directions
As the COVID-19 pandemic evolves, the scientific community’s response will be critical for identifying potential pharmacological and biotechnological developments that may aid in combating the virus and the disease it causes.
However, this global crisis highlights the importance of mounting a response based on collaboration among a wide variety of disciplines.
Understanding the basic science of the virus and its pathogenesis is imperative for identifying and envisioning possible diagnostic and therapeutic approaches; understanding how social factors can influence outcomes and shape implementation of a response is critical to disseminating any scientific advancements.
Summarizing such a complex and ever-changing topic presents a number of challenges.
This review represents the effort of over 50 contributors to distill and interpret the available information.
However, this text represents a dynamic and evolving document, and we welcome continued contributions from all researchers who have insights into how these topics intersect.
A multidisciplinary perspective is critical to understanding this evolving crisis, and in this review we seek to use open science tools to coordinate a response among a variety of researchers.
We intend to publish additional updates as the situation evolves.
9 Discussion
As of October 2020 the SARS-CoV-2 virus remains a serious worldwide threat.
The scientific community has responded by rapidly collecting and disseminating information about the SARS-CoV-2 virus and the associated illness, COVID-19.
The rapid identification of the genomic sequence of the virus allowed for early contextualization of SARS-CoV-2 among other known respiratory viruses.
The pathogen is a coronavirus that is closely related to SARS-CoV-1, which caused the SARS pandemics of the early 2000s.
Knowing the phylogenetic context and genomic sequence of the virus then allowed for rapid insights into its structure and pathogenesis.
As with other HCoV, the immune response to SARS-CoV-2 is likely driven by detection of its spike protein, which allows it to enter cells through the ACE2 receptor.
Epithelial cells have also emerged as the major cellular target of the virus, contextualizing the respiratory and gastrointestinal symptoms that are frequently observed in COVID-19.
However, as COVID-19 cases have been more widely characterized, it has become clear that the disease presentation is highly heterogeneous.
Many cases, especially in younger adults, present with mild symptoms or even asymptomatically, while others, especially in older adults, can be severe or fatal.
In children, the SARS-CoV-2 virus can present as two distinct diseases, COVID-19 or MIS-C.
While the overall infection fatality rate remains unknown, estimates suggest that it is not more than 1%; however, the severity of many non-lethal cases makes COVID-19 an ongoing, significant concern.
Characterizing the rate of infection and fatality rates hinges on the availability of rapid and accurate diagnostic testing.
Major advancements have been made in identifying diagnostic approaches.
The development of diagnostic technologies have been rapid, beginning with the release of the SARS-CoV-2 viral genome sequence in January.
As of October 2020, a range of diagnostic tests have become available.
One class of tests uses PCR (RT-PCR or qRT-PCR) to assess the presence of SARS-CoV-2 RNA, while another typically uses ELISA to test for the presence of antibodies to SARS-CoV-2.
The former approach is useful for identifying active infections, while the latter measures hallmarks of the immune response and therefore can detect either active infections or immunity gained from prior infection.
Combining these tests leads to extremely accurate detection of SARS-CoV-2 infection (98.6%), but when used alone, PCR-based tests are recommended before 5.5 days after the onset of the illness and antibody tests after 5.5 days [400].
Other strategies for testing can also influence the tests’ accuracy, such as the use of nasopharyngeal swabs versus BALF [400], which allow for trade-offs between patient’s comfort and test sensitivity.
Additionally, technologies such as digital PCR may allow for scale-up in the throughput of diagnostic testing, facilitating widespread testing.
One major question that remains is whether people who recover from SARS-CoV-2 develop sustained immunity, and over what period this immunity is expected to last.
Some reports have suggested that some patients may develop COVID-19 reinfections (e.g., [384]), but the rates of reinfection are currently unknown.
Serologic testing combined with PCR testing will be critical to confirming purported cases of reinfection and to identifying the duration over which immunity is retained and to understanding reinfection risks.
One of the goals of characterizing the immune response is to identify strategies for the prophylactic enhancements of immunity.
Though some concerns remain about the duration of sustained immunity for convalescents, vaccine development efforts are ongoing and show initial promising results.
The Moderna trial, for example, reported that the neutralizing activity in participants who received two doses of the vaccine was similar to that observed in convalescent plasma.
Vaccine development for COVID-19 is progressing rapidly compared to typical timelines, but vaccine development is still a lengthy process.
In the meantime, some advances have also been made in the treatment of patients with COVID-19.
As cases have become better characterized, it has become evident that many patients experience an initial immune response to the virus that is typically characterized by fever, cough, dyspnea, and related symptoms.
However, the most serious concern is cytokine release syndrome, when the body’s immune response becomes dysregulated, resulting in an extreme inflammatory response.
The RECOVERY trial, a large-scale, multi-arm trial enrolling about 15% of all COVID-19 patients in the United Kingdom, was the first to identify that the widely available steroid dexamethasone seems to be beneficial for patients suffering from this immune dysregulation [546].
Efforts to identify therapeutic treatments to treat patients early in the course of infection have been more ambiguous.
Early interest in the drugs hydroxychloroquine and chloroquine yielded no promising results from studies with robust experimental designs.
The experimental drug remdesivir, which was developed for Ebola, has received enough support from early analyses to receive FDA approval, but results have been mixed.
The potential for other drugs, such as tocilizumab, to reduce recovery time remains unclear, but some early results were promising.
9.1 Additional Therapeutics of Interest
Given what is currently known about these therapeutics for COVID-19, a number of related therapies beyond those explored above may also prove to be of interest.
For example, the demonstrated benefit of dexamethasone and the ongoing potential of tocilizumab for treatment of COVID-19 suggests that other anti-inflammatory agents might also hold value for the treatment of COVID-19.
Given that current evidence about treating COVID-19 with dexamethasone suggests that the need to curtail the cytokine storm inflammatory response to the virus can transcend the risks of immunosuppression, exploration of more anti-inflammatory agents may be warranted.
While dexamethasone is considered widely available and generally affordable, the high costs of biologics such as tocilizumab therapy may present obstacles to wide-scale distribution of this drug if it proves of value.
At the doses used for rheumatoid arthritis patients, the cost for tocilizumab ranges from $179.20 to $896 per dose for the IV form and $355 for the pre-filled syringe [628].
There are several anti-inflammatory agents used for the treatment of autoimmune diseases that may also be able to counter the effects of the cytokine storm induced by the virus, some of which, such as cyclosporine, are likely to be more cost-effective and readily available than biologics [629].
While tocilizumab targets IL-6, several other inflammatory markers could be potential targets, including TNF-alpha.
Inhibition of TNF-alpha by an inhibitor such as Etanercept has been previously suggested for treatment of SARS-CoV-1 [630] and may be relevant for SARS-CoV-2 as well.
Another anti-IL-6 antibody, sarilumab, is also being investigated [631,632].
Baricitinib and other small molecule inhibitors of the Janus-activated kinase pathway also curtail the inflammatory response and have been suggested as potential options for SARS-CoV-2 infections [633].
Baricitinib in particular may be able to reduce the ability of SARS-CoV-2 to infect lung cells [634].
Clinical trials studying baricitinib in COVID-19 have already begun in the US and in Italy [635,636].
Identification and targeting of further inflammatory markers that are relevant in SARS-CoV-2 infection may be of value for curtailing the inflammatory response and lung damage.
In addition to immunosuppressive treatments that are most beneficial late in disease progression, much research is focused on identifying treatments would be likely to benefit early-stage patients.
For example, although studies of hydroxychloroquine have not supported the early theory-driven interest in this antiviral treatment, alternative compounds with related mechanisms may still have potential.
Hydroxyferroquine derivatives of HCQ have been described as a class of bioorganometallic compounds that exert antiviral effects with some selectivity for SARS-CoV-1 in vitro[637].
Future work could explore whether such compounds exert antiviral effects against SARS-CoV-2 and whether they would be safer for use in COVID-19.
Another potential approach is the development of antivirals, which could be broad-spectrum, specific to coronaviruses, or targeted to SARS-CoV-2.
Development of new antivirals is complicated by the fact that none have yet been approved for human coronaviruses.
Intriguing new options are emerging, however.
Beta-D-N4-hydroxycytidine (NHC) is an orally bioavailable ribonucleotide analog showing broad-spectrum activity against RNA viruses, which may inhibit SARS-CoV-2 replication in vitro and in vivo in mouse models of HCoVs [638].
A range of other antivirals are also in development.
Development of antivirals will be further facilitated as research reveals more information about the interaction of SARS-CoV-2 with the host cell and host cell genome, mechanisms of viral replication, mechanisms of viral assembly, and mechanisms of viral release to other cells; this can allow researchers to target specific stages and structures of the viral life cycle.
Many researchers have also focused their attention to the potential use of dietary supplements and nutraceuticals.
Indeed, there has been recent interest for the potential use of vitamin D as a prophylactic and therapeutic agent against COVID-19 as several observational studies have linked low vitamin D status to its incidence [770,778].
These associations have yet to be confirmed and rigorous trials are required before considering supplementation recommendations.
However, the nutritional status and general health of a patient can affect their outcomes in various diseases, thus it would be pertinent to advise people to follow a healthy diet and life style to the best of their ability to prevent nutrient deficiencies and insufficiencies and to maintain a healthy immune system [644].
Finally, antibodies against viruses, also known as antiviral monoclonal antibodies, could be an alternative as well and are described in detail in an above section.
The goal of antiviral antibodies is to neutralize viruses through either cell-killing activity or blocking of viral replication [639].
They may also engage the host immune response, encouraging the immune system to hone in on the virus.
Given the cytokine storm that results from immune system activation in response to the virus, which has been implicated in worsening of the disease, a neutralizing antibody (nAb) may be preferable.
Upcoming work may explore the specificity of nAbs for their target, mechanisms by which the nAbs impede the virus, and improvements to antibody structure that may enhance the ability of the antibody to block viral activity.
Some research is also investigating potential therapeutics and prophylactics that would interact with components of the innate immune response.
For example, there are a variety of TLRs, PRRs that recognize PAMPs and DAMPs.
TLRs form a part of innate immune recognition and can more generally contribute to promoting both innate and adaptive responses [144].
In mouse models, poly(I:C) and CpG, which are agonists of toll-like receptors TLR3 and TLR9, respectively, showed protective effects when administered prior to SARS-CoV-1 infection [640].
Therefore, TLR agonists hold some potential for broad-spectrum prophylaxis.
Given that a large number of clinical trials are currently in progress, more information about the potential of these and other therapeutics should become available over time.
This information, combined with advances in understanding the molecular structure and viral pathogenesis of SARS-CoV-2, may lead to a more complete understanding of how the virus affects the human host and what strategies can improve outcomes.
To date, investigations of potential therapeutics for COVID-19 have focused primarily on repurposing existing drugs.
This approach is necessary given the urgency of the situation as well as the extensive time required for developing and testing new therapies.
However, in the long-term, new drugs specific for treatment of COVID-19 may also enter development.
Development of novel drugs is likely to be guided by what is known about the pathogenesis and molecular structure of SARS-CoV-2.
For example, understanding the various structural components of SARS-CoV-2 may allow for the development of small molecule inhibitors of those components.
Currently, crystal structures of the SARS-CoV-2 main protease have recently been resolved [459,641], and efforts are already in place to perform screens for small molecule inhibitors of the main protease, which have yielded potential hits [459].
Much work remains to be done to determine further crystal structures of other viral components, understand the relative utility of targeting different viral components, perform additional small molecule inhibitor screens, and determine the safety and efficacy of the potential inhibitors.
While still nascent, work in this area is promising.
Over the longer term, this approach and others may lead to the development of novel therapeutics specifically for COVID-19 and SARS-CoV-2.
10 Application of an Open Publishing Framework to an Emerging Public health Crisis
10.1 Abstract
10.2 Introduction
As international attention remains focused on the ongoing public health crisis, the scientific community has responded by mobilizing resources and turning much of its attention to the virus and disease.
This rapid influx of information is disseminated by traditional publishing mechanisms, preprint servers, and press releases, which provide a venue for scientists to release findings without undergoing the formal publication process.
While having information available is valuable to efforts to understand and combat COVID-19, many contributions come from researchers across a wide range of fields who have varying degrees of experience working on coronaviruses and related topics.
The volume of information available, much of which has not gone through rigorous peer review, presents a significant challenge to individual efforts to keep abreast of the state of COVID-19 research [8].
However, research on these topics is proceeding so rapidly that any static review is likely to quickly become dated.
Our goal as a community is to consolidate information about the virus in the context of related viruses and to synthesize rapidly emerging literature centered on the diagnosis and treatment of COVID-19.
We used an open publishing framework, Manubot [9], to manage hundreds of contributions from the community to create a living, scholarly document.
We designed software to generate figures that automatically update using external data sources.
Our primary goal is to sort and distill informative content out of the overwhelming flood of information [8] and help the broader scientific community become more conversant on this critical subject.
Thus, our approach has been to develop a real-time, collaborative effort that welcomes submissions from scientists worldwide into this ongoing effort.
This document represents the first snapshot, which aims to reflect the state of the field as of October, 2020.
We plan to refine and expand this document until technologies to mitigate the pandemic are widely available.
In an effort to keep pace as new information about COVID-19 and SARS-CoV-2 becomes available, this project is an open, collaborative effort that invited contributions from the scientific community broadly, similar to previous efforts to develop collaborative reviews [1239,1240].
Contributors were recruited by word of mouth and on Twitter.
Existing efforts to train early-career scientists were also integrated: Appendix A contains summaries written by the students, post-docs, and faculty of the Immunology Institute at the Mount Sinai School of Medicine [1241,1242], and two of the authors were recruited through the American Physician Scientist Association’s Virtual Summer Research Program [1243].
The project was managed through GitHub [1244] using Manubot [9] to continuously generate a version of the manuscript online [1245].
Contributors developed text that was proposed through GitHub’s pull request system and then reviewed and approved by at least one other author.
While this document reflects the current version of record, the online version will continue to be developed as information about the pandemic emerges.
Below, we will describe the processes used to synthesize the literature.
10.3 Technical Infrastructure
10.3.1 Collaborative Writing and Manuscript Generation
Manubot [9] is a collaborative framework developed to adapt open-source software development techniques and version control for manuscript writing.
Here, Manubot was used to generate a manuscript from text maintained using GitHub, a popular, online version control interface.
The GitHub implementation allowed users to contribute either using git on the command line or using the GitHub user interface, and we developed documentation for users with less experience with this platform.
Manubot also provides a functionality to create a bibliography using digital object identifiers (DOIs), website URLs, or other identifiers such as PubMed identifiers and arXiv IDs.
Due to the needs of this project, project contributors also implemented new features in Manubot and Zotero, which Manubot uses to extract metadata for some types of citations.
These features support directly citing clinical trial identifiers such as clinicaltrials:NCT04292899[433] and generating the complete review manuscript along with the individual manuscripts reviewing specific topics.
Finally, Manubot and GitHub Actions continuous integration allowed for scripted updates to be run each time the manuscript was generated.
These scripts were used to check that the manuscript was built correctly, run spellchecking, and cross-reference the manuscripts cited in this review, summarized in Appendix A, and discussed in the project’s issues and pull requests.
10.3.2 Data Analysis and Visualization
The combination of Manubot and GitHub Actions also made it possible to dynamically update information such as statistics and visualizations in the manuscript.
Data about worldwide cases and deaths from the COVID-19 Data Repository by the Center for Systems Science and Engineering at Johns Hopkins University [402] were read using a Python script.
Similarly, the clinical trials statistics and figure were generated based on data from the University of Oxford Evidence-Based Medicine Data Lab’s COVID-19 TrialsTracker [405].
In both cases, frequency data were plotted using Matplotlib [1246] in Python.
The figure showing the geographic distribution of COVID-19 clinical trials was generated using the countries associated with the trials listed in the COVID-19 TrialsTracker, converting the country names to 3-letter ISO codes using pycountry or manual adjustment when necessary, and visualizing the geographic distribution of trial recruitment using geopandas.
GitHub Actions runs a nightly workflow to update these external data and regenerate the statistics and figures for the manuscript.
The workflow uses the GitHub API to detect and save the latest commit of the external data sources, which are both GitHub repositories.
It then downloads versioned data from that snapshot of the external repositories and runs bash and Python scripts to calculate the desired statistics and produce the summary figures.
The statistics are stored in JSON files that are accessed by Manubot to populate the values of placeholder template variables dynamically every time the manuscript is built.
For instance, the template variable {{ebm_trials_results}} in the manuscript is replaced by the actual number of clinical trials with results, 98.
The template variables also include versioned URLs to the dynamically updated figures.
The JSON files and figures are stored in the external-resources branch of the manuscript’s GitHub repository, which acts as versioned storage.
The GitHub Actions workflow automatically adds and commits the new JSON files and figures to the external-resources branch every time it runs, and Manubot uses the latest version of these resources when it builds the manuscript.
Relevant articles were identified and submitted as issues on GitHub for review.
Articles were classified as diagnostic, therapeutic, or other, and a template was developed to guide the review of papers and preprints in each category.
Following a framework often used for assessing medical literature, the review consisted of examining methods used in each relevant article, assignment (whether the study was observational or randomized), assessment, results, interpretation, and how well the study extrapolates [1247].
For examples of each template, please see Appendices B-D.
10.4.1 Diagnostic Papers
10.4.1.1 Methods
Reviewers began by describing the study question(s) being investigated by the article.
They then described the study population, the sample size, the prevalence of the disease in the study population, countries / regions considered in case of human subjects, demographics of participants, the setting, and any remaining inclusion / exclusion criteria considered.
They then described the reference test or “gold standard,” if one was utilized.
10.4.1.2 Assignment
Reviewers described how new and reference tests were assigned, including additional relevant details about the study design.
For example, reviewers were asked whether the diagnostic test resulted in rigorous assignments of case status or was biased towards sicker or healthier individuals.
10.4.1.3 Assessment
Reviewers described how the test was performed.
For example, for both standard and reference tests, reviewers described technical details of assays used, when measurements were taken and by whom.
Subsequently, they described how individuals were classified as positive or negative cases and whether results were precise and reproducible with repeated tests.
Reviewers described whether there were any missing data, whether some participants underwent only one test, or whether there were individuals with inconclusive results.
10.4.1.4 Results
Reviewers reported the estimated sensitivity, specificity, positive predictive value (PPV), and negative predicted value (NPV), as well as confidence bounds around these measures, if provided.
10.4.1.5 Interpretation
Reviewers reported how well the test ruled in or ruled out disease based on the population, if there were identified side effects, and patient adherence.
10.4.1.6 Extrapolation
Reviewers described how well this test will extrapolate outside the measured population.
10.4.2 Therapeutic Papers
10.4.2.1 Methods
Reviewers began by describing the study question(s) being investigated by the article.
They then described the study population, the sample size, the prevalence of the disease in the study population, countries / regions considered in case of human subjects, demographics of participants, the setting, and any remaining inclusion / exclusion criteria considered.
10.4.2.2 Assignment
Reviewers described how the treatment is assigned, whether it was an interventional or observational study, whether randomization took place, etc.
10.4.2.3 Assessment
10.4.2.3.1 Outcome Assessment
Reviewers described the outcome that was assessed and evaluated whether it was appropriate given the underlying study question.
They described whether there were any missing data such as whether there were individuals lost to follow up.
They then describe whether there were any potential sources of bias such as lack of blinding in a randomized controlled trial.
10.4.2.3.2 Statistical Methods Assessment
Reviewers described which statistical methods were used for inference and whether applied methods were appropriate for the study.
They then described whether adjustments were made for possible confounders.
10.4.2.4 Results
Reviewers described the estimated association between the treatment and outcome.
They described measures of confidence or statistical significance, if provided.
10.4.2.5 Interpretation
Reviewers described whether a causal claim could be made.
They described whether any side effects or interactions with other drugs were identified, as well as any subgroup findings.
10.4.2.6 Extrapolation
Reviewers describe how the study may extrapolate to a different species or population.
10.5 Conclusions
Figure 8:Summary of the relationships among topics covered in this review.
Several review articles on aspects of COVID-19 have already been published.
These have included reviews on the disease epidemiology [41], immunological response [42], diagnostics [43], and pharmacological treatments [42,44].
Others [45,46] provide narrative reviews of progress on some important ongoing COVID-19 research questions.
With the worldwide scientific community uniting during 2020 to investigate SARS-CoV-2 and COVID-19 from a wide range of perspectives, findings from many disciplines are relevant on a rapid timescale to a broad scientific audience.
Additionally, many findings are published as preprints, which are available prior to going through the peer review process.
As a result, centralizing, summarizing, and critiquing new literature broadly relevant to COVID-19 can help to expedite the interdisciplinary scientific process that is currently happening at an advanced pace.
We are particularly interested in providing background to the development of diagnostic, prophylactic, and therapeutic approaches to COVID-19.
Two major concerns within diagnosis include the detection of current infections in individuals with and without symptoms, and the detection of past exposure without an active infection.
In the latter category, identifying whether individuals can develop or have developed sustained immunity is also a major consideration.
The development of high-throughput, affordable methods for detecting active infections and sustained immunity will be critical to understanding and controlling the disease.
The identification of interventions that can mitigate the effect of the virus on exposed and infected individuals is a significant research priority.
Some possible approaches include the identification of existing pharmaceuticals that reduce the severity of infection, either by reducing the virus’ virulence (e.g., antivirals) or managing the most severe symptoms of infection.
Due to the long timeline for the development of novel pharmaceuticals, in most cases, research surrounding possible pharmaceutical interventions focuses on the identification and investigation of existing compounds whose mechanisms may be relevant to COVID-19.
Other foci of current research include the identification of antibodies produced by survivors of COVID-19 and the development of vaccines.
Understanding the mechanisms describing host-virus interactions between humans and SARS-CoV-2 is thus critical to identifying candidate therapeutics.
An overview of the topics covered is visualized in Figure 8.
Thus, in this review, we seek to consolidate information about efforts to develop strategies for diagnosis and therapeutics as new information is released by the scientific community.
We include information from both traditional peer-reviewed scientific literature and from preprints, which typically have not undergone peer review but have been critically evaluated by the scientists involved in this effort.
The goal of this manuscript is to present preliminary findings within the broader context of COVID-19 research and to identify the broad interpretations of new research, as well as limitations to interpretability.
11 Additional Items
11.1 Competing Interests
Author
Competing Interests
Last Reviewed
Halie M. Rando
None
2021-01-20
Casey S. Greene
None
2021-01-20
Michael P. Robson
None
2020-11-12
Simina M. Boca
None
2020-11-07
Nils Wellhausen
None
2020-11-03
Ronan Lordan
None
2020-11-03
Christian Brueffer
Employee and shareholder of SAGA Diagnostics AB.
2020-11-11
Sandipan Ray
None
2020-11-11
Lucy D'Agostino McGowan
Received consulting fees from Acelity and Sanofi in the past five years
2020-11-10
Anthony Gitter
Filed a patent application with the Wisconsin Alumni Research Foundation related to classifying activated T cells
2020-11-10
Anna Ada Dattoli
None
2020-03-26
Ryan Velazquez
None
2020-11-10
John P. Barton
None
2020-11-11
Jeffrey M. Field
None
2020-11-12
Bharath Ramsundar
None
2020-11-11
Adam L. MacLean
None
2021-02-23
Alexandra J. Lee
None
2020-11-09
Immunology Institute of the Icahn School of Medicine
None
2020-04-07
Fengling Hu
None
2020-04-08
Nafisa M. Jadavji
None
2020-11-11
Elizabeth Sell
None
2020-11-11
Jinhui Wang
None
2021-01-21
Diane N. Rafizadeh
None
2020-11-11
Ashwin N. Skelly
None
2020-11-11
Marouen Ben Guebila
None
2020-11-11
Likhitha Kolla
None
2020-11-16
David Manheim
None
2020-11-11
Soumita Ghosh
None
2020-11-09
James Brian Byrd
Funded by FastGrants to conduct a COVID-19-related clinical trial
2020-11-12
YoSon Park
Now employed by Pfizer (subsequent to contributions to this project)
2020-01-22
Vikas Bansal
None
2021-01-25
Stephen Capone
None
2020-11-11
John J. Dziak
None
2020-11-11
Yuchen Sun
None
2020-11-11
Yanjun Qi
None
2020-07-09
Lamonica Shinholster
None
2020-11-11
Temitayo Lukan
None
2020-11-10
Sergey Knyazev
None
2020-11-11
Dimitri Perrin
None
2020-11-11
Serghei Mangul
None
2020-11-11
Shikta Das
None
2020-08-13
Gregory L Szeto
None
2020-11-16
Tiago Lubiana
None
2020-11-11
David Mai
None
2021-01-08
COVID-19 Review Consortium
None
2021-01-16
Rishi Raj Goel
None
2021-01-20
Joel D Boerckel
None
2021-03-26
11.2 Author Contributions
Author
Contributions
Halie M. Rando
A, D, E, Project Administration, Writing - Original Draft, Writing - Review & Editing
Writing - Original Draft, Writing - Review & Editing
Marouen Ben Guebila
Writing - Original Draft, Writing - Review & Editing
Likhitha Kolla
Writing - Original Draft
David Manheim
Investigation, Writing - Original Draft
Soumita Ghosh
Writing - Original Draft
James Brian Byrd
Writing - Original Draft, Writing - Review & Editing
YoSon Park
Writing - Original Draft, Writing - Review & Editing
Vikas Bansal
Writing - Original Draft, Writing - Review & Editing
Stephen Capone
Writing - Original Draft, Writing - Review & Editing
John J. Dziak
Writing - Original Draft, Writing - Review & Editing
Yuchen Sun
Visualization
Yanjun Qi
Visualization
Lamonica Shinholster
Writing - Original Draft
Temitayo Lukan
Investigation, Writing - Original Draft
Sergey Knyazev
Writing - Original Draft, Writing - Review & Editing
Dimitri Perrin
Writing - Original Draft, Writing - Review & Editing
Serghei Mangul
Writing - Review & Editing
Shikta Das
Writing - Review & Editing
Gregory L Szeto
Writing - Review & Editing
Tiago Lubiana
Writing - Review & Editing
David Mai
Writing - Original Draft, Writing - Review & Editing
COVID-19 Review Consortium
Project Administration
Rishi Raj Goel
Writing - Original Draft, Writing - Review & Editing
Joel D Boerckel
Writing - Review & Editing
11.3 Acknowledgements
We thank Nick DeVito for assistance with the Evidence-Based Medicine Data Lab COVID-19 TrialsTracker data and Vincent Rubinetti and Daniel Himmelstein for feedback on and support with Manubot.
We thank Yael Evelyn Marshall who contributed writing (original draft) as well as reviewing and editing of pieces of the text but who did not formally approve the manuscript, as well as Ronnie Russell, who contributed text to and helped develop the structure of the manuscript early in the writing process and Matthias Fax who helped with writing and editing text related to diagnostics.
We are grateful to the following contributors for reviewing pieces of the text: Nadia Danilova, James Eberwine and Ipsita Krishnan.
12 References
1. Pathogenesis, Symptomatology, and Transmission of SARS-CoV-2 through analysis of Viral Genomics and Structure
Halie M. Rando, Adam L. MacLean, Alexandra J. Lee, Sandipan Ray, Vikas Bansal, Ashwin N. Skelly, Elizabeth Sell, John J. Dziak, Lamonica Shinholster, Lucy D’Agostino McGowan, … Casey S. Greene arXiv (2021-02-15) https://arxiv.org/abs/2102.01521
2. Dietary Supplements and Nutraceuticals Under Investigation for COVID-19 Prevention and Treatment
Ronan Lordan, Halie M. Rando, COVID-19 Review Consortium, Casey S. Greene arXiv (2021-02-05) https://arxiv.org/abs/2102.02250
3. Identification and Development of Therapeutics for COVID-19
Halie M. Rando, Nils Wellhausen, Soumita Ghosh, Alexandra J. Lee, Anna Ada Dattoli, Fengling Hu, James Brian Byrd, Diane N. Rafizadeh, Yanjun Qi, Yuchen Sun, … Casey S. Greene arXiv (2021-03-05) https://arxiv.org/abs/2103.02723
9. Open collaborative writing with Manubot
Daniel S. Himmelstein, Vincent Rubinetti, David R. Slochower, Dongbo Hu, Venkat S. Malladi, Casey S. Greene, Anthony Gitter PLOS Computational Biology (2019-06-24) https://doi.org/c7np
DOI: 10.1371/journal.pcbi.1007128 · PMID: 31233491 · PMCID: PMC6611653
12. Managing epidemics: key facts about major deadly diseases.
World Health Organization
(2018)
ISBN: 9789241565530
13. A Novel Coronavirus Genome Identified in a Cluster of Pneumonia Cases — Wuhan, China 2019−2020
Wenjie Tan, Xiang Zhao, Xuejun Ma, Wenling Wang, Peihua Niu, Wenbo Xu, George F. Gao, Guizhen Wu, MHC Key Laboratory of Biosafety, National Institute for Viral Disease Control and Prevention, China CDC, Beijing, China, Center for Biosafety Mega-Science, Chinese Academy of Sciences, Beijing, China China CDC Weekly (2020) https://doi.org/gg8z47
DOI: 10.46234/ccdcw2020.017
14. A new coronavirus associated with human respiratory disease in China
Fan Wu, Su Zhao, Bin Yu, Yan-Mei Chen, Wen Wang, Zhi-Gang Song, Yi Hu, Zhao-Wu Tao, Jun-Hua Tian, Yuan-Yuan Pei, … Yong-Zhen Zhang Nature (2020-02-03) https://doi.org/dk2w
DOI: 10.1038/s41586-020-2008-3 · PMID: 32015508 · PMCID: PMC7094943
15. A pneumonia outbreak associated with a new coronavirus of probable bat origin
Peng Zhou, Xing-Lou Yang, Xian-Guang Wang, Ben Hu, Lei Zhang, Wei Zhang, Hao-Rui Si, Yan Zhu, Bei Li, Chao-Lin Huang, … Zheng-Li Shi Nature (2020-02-03) https://doi.org/ggj5cg
DOI: 10.1038/s41586-020-2012-7 · PMID: 32015507 · PMCID: PMC7095418
16. Genomic characterisation and epidemiology of 2019 novel coronavirus: implications for virus origins and receptor binding
Roujian Lu, Xiang Zhao, Juan Li, Peihua Niu, Bo Yang, Honglong Wu, Wenling Wang, Hao Song, Baoying Huang, Na Zhu, … Wenjie Tan The Lancet (2020-02) https://doi.org/ggjr43
DOI: 10.1016/s0140-6736(20)30251-8
17. On the origin and continuing evolution of SARS-CoV-2
Xiaolu Tang, Changcheng Wu, Xiang Li, Yuhe Song, Xinmin Yao, Xinkai Wu, Yuange Duan, Hong Zhang, Yirong Wang, Zhaohui Qian, … Jian Lu National Science Review (2020-06) https://doi.org/ggndzn
DOI: 10.1093/nsr/nwaa036 · PMCID: PMC7107875
25. Recovery in tracheal organ cultures of novel viruses from patients with respiratory disease.
K. McIntosh, J. H. Dees, W. B. Becker, A. Z. Kapikian, R. M. Chanock Proceedings of the National Academy of Sciences (1967-04-01) https://doi.org/bhfd6w
DOI: 10.1073/pnas.57.4.933 · PMID: 5231356 · PMCID: PMC224637
28. Identification of a new human coronavirus
Lia van der Hoek, Krzysztof Pyrc, Maarten F Jebbink, Wilma Vermeulen-Oost, Ron JM Berkhout, Katja C Wolthers, Pauline ME Wertheim-van Dillen, Jos Kaandorp, Joke Spaargaren, Ben Berkhout Nature Medicine (2004-03-21) https://doi.org/b5wtsn
DOI: 10.1038/nm1024 · PMID: 15034574 · PMCID: PMC7095789
29. Characterization and Complete Genome Sequence of a Novel Coronavirus, Coronavirus HKU1, from Patients with Pneumonia
Patrick C. Y. Woo, Susanna K. P. Lau, Chung-ming Chu, Kwok-hung Chan, Hoi-wah Tsoi, Yi Huang, Beatrice H. L. Wong, Rosana W. S. Poon, James J. Cai, Wei-kwang Luk, … Kwok-yung Yuen Journal of Virology (2005-01-15) https://doi.org/bk7m7h
DOI: 10.1128/jvi.79.2.884-895.2005 · PMID: 15613317 · PMCID: PMC538593
30. Coronavirus 229E-Related Pneumonia in Immunocompromised Patients
F. Pene, A. Merlat, A. Vabret, F. Rozenberg, A. Buzyn, F. Dreyfus, A. Cariou, F. Freymuth, P. Lebon Clinical Infectious Diseases (2003-10-01) https://doi.org/dcjk64
DOI: 10.1086/377612 · PMID: 13130404 · PMCID: PMC7107892
31. SARS and MERS: recent insights into emerging coronaviruses
Emmie de Wit, Neeltje van Doremalen, Darryl Falzarano, Vincent J. Munster Nature Reviews Microbiology (2016-06-27) https://doi.org/f8v5cv
DOI: 10.1038/nrmicro.2016.81 · PMID: 27344959 · PMCID: PMC7097822
37. Clinical Characteristics of Coronavirus Disease 2019 in China
Wei-jie Guan, Zheng-yi Ni, Yu Hu, Wen-hua Liang, Chun-quan Ou, Jian-xing He, Lei Liu, Hong Shan, Chun-liang Lei, David S. C. Hui, … Nan-shan Zhong New England Journal of Medicine (2020-04-30) https://doi.org/ggm6dh
DOI: 10.1056/nejmoa2002032 · PMID: 32109013 · PMCID: PMC7092819
38. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: a retrospective cohort study
Fei Zhou, Ting Yu, Ronghui Du, Guohui Fan, Ying Liu, Zhibo Liu, Jie Xiang, Yeming Wang, Bin Song, Xiaoying Gu, … Bin Cao The Lancet (2020-03) https://doi.org/ggnxb3
DOI: 10.1016/s0140-6736(20)30566-3
39. Presenting Characteristics, Comorbidities, and Outcomes Among 5700 Patients Hospitalized With COVID-19 in the New York City Area
Safiya Richardson, Jamie S. Hirsch, Mangala Narasimhan, James M. Crawford, Thomas McGinn, Karina W. Davidson, Douglas P. Barnaby, Lance B. Becker, John D. Chelico, Stuart L. Cohen, … and the Northwell COVID-19 Research Consortium JAMA (2020-05-26) https://doi.org/ggsrkd
DOI: 10.1001/jama.2020.6775 · PMID: 32320003 · PMCID: PMC7177629
40. Clinical Characteristics of Covid-19 in New York City
Parag Goyal, Justin J. Choi, Laura C. Pinheiro, Edward J. Schenck, Ruijun Chen, Assem Jabri, Michael J. Satlin, Thomas R. Campion, Musarrat Nahid, Joanna B. Ringel, … Monika M. Safford New England Journal of Medicine (2020-06-11) https://doi.org/ggtsjc
DOI: 10.1056/nejmc2010419 · PMID: 32302078 · PMCID: PMC7182018
41. COVID-19: Epidemiology, Evolution, and Cross-Disciplinary Perspectives
Jiumeng Sun, Wan-Ting He, Lifang Wang, Alexander Lai, Xiang Ji, Xiaofeng Zhai, Gairu Li, Marc A. Suchard, Jin Tian, Jiyong Zhou, … Shuo Su Trends in Molecular Medicine (2020-05) https://doi.org/ggqgwq
DOI: 10.1016/j.molmed.2020.02.008 · PMID: 32359479 · PMCID: PMC7118693
42. Immunology of COVID-19: Current State of the Science
Nicolas Vabret, Graham J. Britton, Conor Gruber, Samarth Hegde, Joel Kim, Maria Kuksin, Rachel Levantovsky, Louise Malle, Alvaro Moreira, Matthew D. Park, … Uri Laserson Immunity (2020-06) https://doi.org/ggt54g
DOI: 10.1016/j.immuni.2020.05.002 · PMID: 32505227 · PMCID: PMC7200337
44. Pharmacologic Treatments for Coronavirus Disease 2019 (COVID-19)
James M. Sanders, Marguerite L. Monogue, Tomasz Z. Jodlowski, James B. Cutrell JAMA (2020-04-13) https://doi.org/ggr27x
DOI: 10.1001/jama.2020.6019 · PMID: 32282022
46. Pathophysiology, Transmission, Diagnosis, and Treatment of Coronavirus Disease 2019 (COVID-19)
W. Joost Wiersinga, Andrew Rhodes, Allen C. Cheng, Sharon J. Peacock, Hallie C. Prescott JAMA (2020-08-25) https://doi.org/gg4ht4
DOI: 10.1001/jama.2020.12839 · PMID: 32648899
49. Acute lung injury in patients with COVID‐19 infection
Liyang Li, Qihong Huang, Diane C. Wang, David H. Ingbar, Xiangdong Wang Clinical and Translational Medicine (2020-03-31) https://doi.org/ghqcrz
DOI: 10.1002/ctm2.16 · PMID: 32508022 · PMCID: PMC7240840
51. A familial cluster of pneumonia associated with the 2019 novel coronavirus indicating person-to-person transmission: a study of a family cluster
Jasper Fuk-Woo Chan, Shuofeng Yuan, Kin-Hang Kok, Kelvin Kai-Wang To, Hin Chu, Jin Yang, Fanfan Xing, Jieling Liu, Cyril Chik-Yan Yip, Rosana Wing-Shan Poon, … Kwok-Yung Yuen The Lancet (2020-02) https://doi.org/ggjs7j
DOI: 10.1016/s0140-6736(20)30154-9 · PMID: 31986261 · PMCID: PMC7159286
52. Fields virology
Bernard N. Fields, David M. Knipe, Peter M. Howley (editors) Wolters Kluwer Health/Lippincott Williams & Wilkins (2007)
ISBN: 9780781760607
53. Important Role for the Transmembrane Domain of Severe Acute Respiratory Syndrome Coronavirus Spike Protein during Entry
Rene Broer, Bertrand Boson, Willy Spaan, François-Loïc Cosset, Jeroen Corver Journal of Virology (2006-02-01) https://doi.org/dvvg2h
DOI: 10.1128/jvi.80.3.1302-1310.2006 · PMID: 16415007 · PMCID: PMC1346921
54. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation
Daniel Wrapp, Nianshuang Wang, Kizzmekia S. Corbett, Jory A. Goldsmith, Ching-Lin Hsieh, Olubukola Abiona, Barney S. Graham, Jason S. McLellan Science (2020-03-13) https://doi.org/ggmtk2
DOI: 10.1126/science.abb2507 · PMID: 32075877
55. Medical microbiology
Samuel Baron (editor) University of Texas Medical Branch at Galveston (1996)
ISBN: 9780963117212
60. Mechanisms of Coronavirus Cell Entry Mediated by the Viral Spike Protein
Sandrine Belouzard, Jean K. Millet, Beth N. Licitra, Gary R. Whittaker Viruses (2012-06-20) https://doi.org/gbbktb
DOI: 10.3390/v4061011 · PMID: 22816037 · PMCID: PMC3397359
61. Phylogenetic Analysis and Structural Modeling of SARS-CoV-2 Spike Protein Reveals an Evolutionary Distinct and Proteolytically Sensitive Activation Loop
Javier A. Jaimes, Nicole M. André, Joshua S. Chappie, Jean K. Millet, Gary R. Whittaker Journal of Molecular Biology (2020-05) https://doi.org/ggtxhr
DOI: 10.1016/j.jmb.2020.04.009 · PMID: 32320687 · PMCID: PMC7166309
62. Angiotensin-converting enzyme 2 (ACE2) as a SARS-CoV-2 receptor: molecular mechanisms and potential therapeutic target
Haibo Zhang, Josef M. Penninger, Yimin Li, Nanshan Zhong, Arthur S. Slutsky Intensive Care Medicine (2020-03-03) https://doi.org/ggpx6p
DOI: 10.1007/s00134-020-05985-9 · PMID: 32125455 · PMCID: PMC7079879
63. Infection of Human Airway Epithelia by Sars Coronavirus is Associated with ACE2 Expression and Localization
Hong Peng Jia, Dwight C. Look, Melissa Hickey, Lei Shi, Lecia Pewe, Jason Netland, Michael Farzan, Christine Wohlford-Lenane, Stanley Perlman, Paul B. McCray Advances in Experimental Medicine and Biology (2006) https://doi.org/dhh5tp
DOI: 10.1007/978-0-387-33012-9_85 · PMID: 17037581 · PMCID: PMC7123641
64. The protein expression profile of ACE2 in human tissues
Feria Hikmet, Loren Méar, Åsa Edvinsson, Patrick Micke, Mathias Uhlén, Cecilia Lindskog Molecular Systems Biology (2020-07-26) https://doi.org/gg6mxv
DOI: 10.15252/msb.20209610 · PMID: 32715618 · PMCID: PMC7383091
66. The spike protein of SARS-CoV — a target for vaccine and therapeutic development
Lanying Du, Yuxian He, Yusen Zhou, Shuwen Liu, Bo-Jian Zheng, Shibo Jiang Nature Reviews Microbiology (2009-02-09) https://doi.org/d4tq4t
DOI: 10.1038/nrmicro2090 · PMID: 19198616 · PMCID: PMC2750777
67. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein
Alexandra C. Walls, Young-Jun Park, M. Alejandra Tortorici, Abigail Wall, Andrew T. McGuire, David Veesler Cell (2020-04) https://doi.org/dpvh
DOI: 10.1016/j.cell.2020.02.058 · PMID: 32155444 · PMCID: PMC7102599
69. Coronavirus membrane fusion mechanism offers a potential target for antiviral development
Tiffany Tang, Miya Bidon, Javier A. Jaimes, Gary R. Whittaker, Susan Daniel Antiviral Research (2020-06) https://doi.org/ggr23b
DOI: 10.1016/j.antiviral.2020.104792 · PMID: 32272173 · PMCID: PMC7194977
70. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor
Markus Hoffmann, Hannah Kleine-Weber, Simon Schroeder, Nadine Krüger, Tanja Herrler, Sandra Erichsen, Tobias S. Schiergens, Georg Herrler, Nai-Huei Wu, Andreas Nitsche, … Stefan Pöhlmann Cell (2020-04) https://doi.org/ggnq74
DOI: 10.1016/j.cell.2020.02.052 · PMID: 32142651 · PMCID: PMC7102627
71. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV
Xiuyuan Ou, Yan Liu, Xiaobo Lei, Pei Li, Dan Mi, Lili Ren, Li Guo, Ruixuan Guo, Ting Chen, Jiaxin Hu, … Zhaohui Qian Nature Communications (2020-03-27) https://doi.org/ggqsrf
DOI: 10.1038/s41467-020-15562-9 · PMID: 32221306 · PMCID: PMC7100515
75. The endothelial glycocalyx: composition, functions, and visualization
Sietze Reitsma, Dick W. Slaaf, Hans Vink, Marc A. M. J. van Zandvoort, Mirjam G. A. oude Egbrink Pflügers Archiv - European Journal of Physiology (2007-01-26) https://doi.org/fsxzdn
DOI: 10.1007/s00424-007-0212-8 · PMID: 17256154 · PMCID: PMC1915585
76. SARS-CoV-2 Infection Depends on Cellular Heparan Sulfate and ACE2
Thomas Mandel Clausen, Daniel R. Sandoval, Charlotte B. Spliid, Jessica Pihl, Hailee R. Perrett, Chelsea D. Painter, Anoop Narayanan, Sydney A. Majowicz, Elizabeth M. Kwong, Rachael N. McVicar, … Jeffrey D. Esko Cell (2020-11) https://doi.org/ghj58m
DOI: 10.1016/j.cell.2020.09.033 · PMID: 32970989 · PMCID: PMC7489987
77. Role of heparan sulfate in immune system-blood vessel interactions
Nathan S. Ihrcke, Lucile E. Wrenshall, Bonnie J. Lindman, Jeffrey L. Platt Immunology Today (1993-10) https://doi.org/bp82dx
DOI: 10.1016/0167-5699(93)90265-m
78. Heparan sulfate proteoglycans as attachment factor for SARS-CoV-2
Lin Liu, Pradeep Chopra, Xiuru Li, Kim M. Bouwman, S. Mark Tompkins, Margreet A. Wolfert, Robert P. de Vries, Geert-Jan Boons Cold Spring Harbor Laboratory (2021-01-04) https://doi.org/gjgqbf
DOI: 10.1101/2020.05.10.087288 · PMID: 32511404 · PMCID: PMC7263551
80. The Pulmonary Endothelial Glycocalyx in ARDS: A Critical Role for Heparan Sulfate
Wells B. LaRivière, Eric P. Schmidt Current Topics in Membranes (2018) https://doi.org/gjgp98
DOI: 10.1016/bs.ctm.2018.08.005 · PMID: 30360782
81. Heparan sulfate assists SARS-CoV-2 in cell entry and can be targeted by approved drugs in vitro
Qi Zhang, Catherine Zhengzheng Chen, Manju Swaroop, Miao Xu, Lihui Wang, Juhyung Lee, Amy Qiu Wang, Manisha Pradhan, Natalie Hagen, Lu Chen, … Yihong Ye Cell Discovery (2020-11-04) https://doi.org/gjgqbd
DOI: 10.1038/s41421-020-00222-5 · PMID: 33298900 · PMCID: PMC7610239
82. Beneficial non-anticoagulant mechanisms underlying heparin treatment of COVID-19 patients
Baranca Buijsers, Cansu Yanginlar, Marissa L. Maciej-Hulme, Quirijn de Mast, Johan van der Vlag EBioMedicine (2020-09) https://doi.org/gjgqbb
DOI: 10.1016/j.ebiom.2020.102969 · PMID: 32853989 · PMCID: PMC7445140
83. The Effect of Anticoagulation Use on Mortality in COVID-19 Infection
Husam M. Salah, Jwan A. Naser, Giuseppe Calcaterra, Pier Paolo Bassareo, Jawahar L. Mehta The American Journal of Cardiology (2020-11) https://doi.org/gjgp99
DOI: 10.1016/j.amjcard.2020.08.005 · PMID: 32892991 · PMCID: PMC7428681
84. Protective Effects of Lactoferrin against SARS-CoV-2 Infection In Vitro
Claudio Salaris, Melania Scarpa, Marina Elli, Alice Bertolini, Simone Guglielmetti, Fabrizio Pregliasco, Corrado Blandizzi, Paola Brun, Ignazio Castagliuolo Nutrients (2021-01-23) https://doi.org/gjgqbg
DOI: 10.3390/nu13020328 · PMID: 33498631 · PMCID: PMC7911668
85. Glycocalyx as Possible Limiting Factor in COVID-19
Patricia P. Wadowski, Bernd Jilma, Christoph W. Kopp, Sebastian Ertl, Thomas Gremmel, Renate Koppensteiner Frontiers in Immunology (2021-02-22) https://doi.org//10.3389/fimmu.2021.607306
DOI: /10.3389/fimmu.2021.607306
86. Spatiotemporal interplay of severe acute respiratory syndrome coronavirus and respiratory mucosal cells drives viral dissemination in rhesus macaques
L Liu, Q Wei, K Nishiura, J Peng, H Wang, C Midkiff, X Alvarez, C Qin, A Lackner, Z Chen Mucosal Immunology (2015-12-09) https://doi.org/f8r7dk
DOI: 10.1038/mi.2015.127 · PMID: 26647718 · PMCID: PMC4900951
87. Understanding Viral dsRNA-Mediated Innate Immune Responses at the Cellular Level Using a Rainbow Trout Model
Sarah J. Poynter, Stephanie J. DeWitte-Orr Frontiers in Immunology (2018-04-23) https://doi.org/gdhpbs
DOI: 10.3389/fimmu.2018.00829 · PMID: 29740439 · PMCID: PMC5924774
88. Ultrastructure and Origin of Membrane Vesicles Associated with the Severe Acute Respiratory Syndrome Coronavirus Replication Complex
Eric J. Snijder, Yvonne van der Meer, Jessika Zevenhoven-Dobbe, Jos J. M. Onderwater, Jannes van der Meulen, Henk K. Koerten, A. Mieke Mommaas Journal of Virology (2006-06-15) https://doi.org/b2rh4r
DOI: 10.1128/jvi.02501-05 · PMID: 16731931 · PMCID: PMC1472606
89. Structural basis for translational shutdown and immune evasion by the Nsp1 protein of SARS-CoV-2
Matthias Thoms, Robert Buschauer, Michael Ameismeier, Lennart Koepke, Timo Denk, Maximilian Hirschenberger, Hanna Kratzat, Manuel Hayn, Timur Mackens-Kiani, Jingdong Cheng, … Roland Beckmann Science (2020-09-04) https://doi.org/gg69nq
DOI: 10.1126/science.abc8665 · PMID: 32680882 · PMCID: PMC7402621
90. Immune responses in COVID-19 and potential vaccines: Lessons learned from SARS and MERS epidemic
Asian Pacific Journal of Allergy and Immunology
(2020) https://doi.org/ggpvxw
DOI: 10.12932/ap-200220-0772 · PMID: 32105090
91. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus
Wenhui Li, Michael J. Moore, Natalya Vasilieva, Jianhua Sui, Swee Kee Wong, Michael A. Berne, Mohan Somasundaran, John L. Sullivan, Katherine Luzuriaga, Thomas C. Greenough, … Michael Farzan Nature (2003-11) https://doi.org/bqvpjh
DOI: 10.1038/nature02145 · PMID: 14647384 · PMCID: PMC7095016
92. Efficient Activation of the Severe Acute Respiratory Syndrome Coronavirus Spike Protein by the Transmembrane Protease TMPRSS2
Shutoku Matsuyama, Noriyo Nagata, Kazuya Shirato, Miyuki Kawase, Makoto Takeda, Fumihiro Taguchi Journal of Virology (2010-12-15) https://doi.org/d4hnfr
DOI: 10.1128/jvi.01542-10 · PMID: 20926566 · PMCID: PMC3004351
93. Evidence that TMPRSS2 Activates the Severe Acute Respiratory Syndrome Coronavirus Spike Protein for Membrane Fusion and Reduces Viral Control by the Humoral Immune Response
I. Glowacka, S. Bertram, M. A. Muller, P. Allen, E. Soilleux, S. Pfefferle, I. Steffen, T. S. Tsegaye, Y. He, K. Gnirss, … S. Pohlmann Journal of Virology (2011-02-16) https://doi.org/bg97wb
DOI: 10.1128/jvi.02232-10 · PMID: 21325420 · PMCID: PMC3126222
94. Increasing Host Cellular Receptor—Angiotensin-Converting Enzyme 2 (ACE2) Expression by Coronavirus may Facilitate 2019-nCoV Infection
Pei-Hui Wang, Yun Cheng Cold Spring Harbor Laboratory (2020-02-27) https://doi.org/ggscwd
DOI: 10.1101/2020.02.24.963348
96. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis
I Hamming, W Timens, MLC Bulthuis, AT Lely, GJ Navis, H van Goor The Journal of Pathology (2004-06) https://doi.org/bhpzc3
DOI: 10.1002/path.1570 · PMID: 15141377
97. Detection of SARS-CoV-2 in Different Types of Clinical Specimens
Wenling Wang, Yanli Xu, Ruqin Gao, Roujian Lu, Kai Han, Guizhen Wu, Wenjie Tan JAMA (2020-03-11) https://doi.org/ggpp6h
DOI: 10.1001/jama.2020.3786 · PMID: 32159775 · PMCID: PMC7066521
98. Epidemiologic Features and Clinical Course of Patients Infected With SARS-CoV-2 in Singapore
Barnaby Edward Young, Sean Wei Xiang Ong, Shirin Kalimuddin, Jenny G. Low, Seow Yen Tan, Jiashen Loh, Oon-Tek Ng, Kalisvar Marimuthu, Li Wei Ang, Tze Minn Mak, … for the Singapore 2019 Novel Coronavirus Outbreak Research Team JAMA (2020-04-21) https://doi.org/ggnb37
DOI: 10.1001/jama.2020.3204 · PMID: 32125362 · PMCID: PMC7054855
99. Persistence and clearance of viral RNA in 2019 novel coronavirus disease rehabilitation patients
Yun Ling, Shui-Bao Xu, Yi-Xiao Lin, Di Tian, Zhao-Qin Zhu, Fa-Hui Dai, Fan Wu, Zhi-Gang Song, Wei Huang, Jun Chen, … Hong-Zhou Lu Chinese Medical Journal (2020-05-05) https://doi.org/ggnnz8
DOI: 10.1097/cm9.0000000000000774 · PMID: 32118639 · PMCID: PMC7147278
100. Postmortem examination of COVID‐19 patients reveals diffuse alveolar damage with severe capillary congestion and variegated findings in lungs and other organs suggesting vascular dysfunction
Thomas Menter, Jasmin D Haslbauer, Ronny Nienhold, Spasenija Savic, Helmut Hopfer, Nikolaus Deigendesch, Stephan Frank, Daniel Turek, Niels Willi, Hans Pargger, … Alexandar Tzankov Histopathology (2020-07-05) https://doi.org/ggwr32
DOI: 10.1111/his.14134 · PMID: 32364264 · PMCID: PMC7496150
101. Association of Cardiac Injury With Mortality in Hospitalized Patients With COVID-19 in Wuhan, China
Shaobo Shi, Mu Qin, Bo Shen, Yuli Cai, Tao Liu, Fan Yang, Wei Gong, Xu Liu, Jinjun Liang, Qinyan Zhao, … Congxin Huang JAMA Cardiology (2020-07-01) https://doi.org/ggq8qf
DOI: 10.1001/jamacardio.2020.0950 · PMID: 32211816 · PMCID: PMC7097841
107. Symptom Profiles of a Convenience Sample of Patients with COVID-19 — United States, January–April 2020
Rachel M. Burke, Marie E. Killerby, Suzanne Newton, Candace E. Ashworth, Abby L. Berns, Skyler Brennan, Jonathan M. Bressler, Erica Bye, Richard Crawford, Laurel Harduar Morano, … Case Investigation Form Working Group MMWR. Morbidity and Mortality Weekly Report (2020-07-17) https://doi.org/gg8r2m
DOI: 10.15585/mmwr.mm6928a2 · PMID: 32673296 · PMCID: PMC7366851
108. Population-scale Longitudinal Mapping of COVID-19 Symptoms, Behavior, and Testing Identifies Contributors to Continued Disease Spread in the United States
William E. Allen, Han Altae-Tran, James Briggs, Xin Jin, Glen McGee, Andy Shi, Rumya Raghavan, Mireille Kamariza, Nicole Nova, Albert Pereta, … Xihong Lin Cold Spring Harbor Laboratory (2020-06-11) https://doi.org/gg8skn
DOI: 10.1101/2020.06.09.20126813 · PMID: 32577674 · PMCID: PMC7302230
109. Extrapulmonary manifestations of COVID-19
Aakriti Gupta, Mahesh V. Madhavan, Kartik Sehgal, Nandini Nair, Shiwani Mahajan, Tejasav S. Sehrawat, Behnood Bikdeli, Neha Ahluwalia, John C. Ausiello, Elaine Y. Wan, … Donald W. Landry Nature Medicine (2020-07-10) https://doi.org/gg4r37
DOI: 10.1038/s41591-020-0968-3 · PMID: 32651579
110. Acute kidney injury in patients hospitalized with COVID-19
Jamie S. Hirsch, Jia H. Ng, Daniel W. Ross, Purva Sharma, Hitesh H. Shah, Richard L. Barnett, Azzour D. Hazzan, Steven Fishbane, Kenar D. Jhaveri, Mersema Abate, … Jia Hwei. Ng Kidney International (2020-07) https://doi.org/ggx24k
DOI: 10.1016/j.kint.2020.05.006 · PMID: 32416116 · PMCID: PMC7229463
111. Nervous system involvement after infection with COVID-19 and other coronaviruses
Yeshun Wu, Xiaolin Xu, Zijun Chen, Jiahao Duan, Kenji Hashimoto, Ling Yang, Cunming Liu, Chun Yang Brain, Behavior, and Immunity (2020-07) https://doi.org/ggq7s2
DOI: 10.1016/j.bbi.2020.03.031 · PMID: 32240762 · PMCID: PMC7146689
112. Neurological associations of COVID-19
Mark A Ellul, Laura Benjamin, Bhagteshwar Singh, Suzannah Lant, Benedict Daniel Michael, Ava Easton, Rachel Kneen, Sylviane Defres, Jim Sejvar, Tom Solomon The Lancet Neurology (2020-09) https://doi.org/d259
DOI: 10.1016/s1474-4422(20)30221-0 · PMID: 32622375 · PMCID: PMC7332267
116. Olfactory transmucosal SARS-CoV-2 invasion as a port of central nervous system entry in individuals with COVID-19
Jenny Meinhardt, Josefine Radke, Carsten Dittmayer, Jonas Franz, Carolina Thomas, Ronja Mothes, Michael Laue, Julia Schneider, Sebastian Brünink, Selina Greuel, … Frank L. Heppner Nature Neuroscience (2020-11-30) https://doi.org/fk46
DOI: 10.1038/s41593-020-00758-5 · PMID: 33257876
117. Large-Vessel Stroke as a Presenting Feature of Covid-19 in the Young
Thomas J. Oxley, J. Mocco, Shahram Majidi, Christopher P. Kellner, Hazem Shoirah, I. Paul Singh, Reade A. De Leacy, Tomoyoshi Shigematsu, Travis R. Ladner, Kurt A. Yaeger, … Johanna T. Fifi New England Journal of Medicine (2020-05-14) https://doi.org/ggtsjg
DOI: 10.1056/nejmc2009787 · PMID: 32343504 · PMCID: PMC7207073
118. Incidence of thrombotic complications in critically ill ICU patients with COVID-19
F. A. Klok, M. J. H. A. Kruip, N. J. M. van der Meer, M. S. Arbous, D. A. M. P. J. Gommers, K. M. Kant, F. H. J. Kaptein, J. van Paassen, M. A. M. Stals, M. V. Huisman, H. Endeman Thrombosis Research (2020-07) https://doi.org/dt2q
DOI: 10.1016/j.thromres.2020.04.013 · PMID: 32291094 · PMCID: PMC7146714
119. Coagulopathy and Antiphospholipid Antibodies in Patients with Covid-19
Yan Zhang, Meng Xiao, Shulan Zhang, Peng Xia, Wei Cao, Wei Jiang, Huan Chen, Xin Ding, Hua Zhao, Hongmin Zhang, … Shuyang Zhang New England Journal of Medicine (2020-04-23) https://doi.org/ggrgz7
DOI: 10.1056/nejmc2007575 · PMID: 32268022 · PMCID: PMC7161262
120. Abnormal coagulation parameters are associated with poor prognosis in patients with novel coronavirus pneumonia
Ning Tang, Dengju Li, Xiong Wang, Ziyong Sun Journal of Thrombosis and Haemostasis (2020-04) https://doi.org/ggqxf6
DOI: 10.1111/jth.14768 · PMID: 32073213 · PMCID: PMC7166509
121. Review: Viral infections and mechanisms of thrombosis and bleeding
M. Goeijenbier, M. van Wissen, C. van de Weg, E. Jong, V. E. A. Gerdes, J. C. M. Meijers, D. P. M. Brandjes, E. C. M. van Gorp Journal of Medical Virology (2012-10) https://doi.org/f37tfr
DOI: 10.1002/jmv.23354 · PMID: 22930518 · PMCID: PMC7166625
122. Immune mechanisms of pulmonary intravascular coagulopathy in COVID-19 pneumonia
Dennis McGonagle, James S O’Donnell, Kassem Sharif, Paul Emery, Charles Bridgewood The Lancet Rheumatology (2020-07) https://doi.org/ggvd74
DOI: 10.1016/s2665-9913(20)30121-1 · PMID: 32835247 · PMCID: PMC7252093
123. “War to the knife” against thromboinflammation to protect endothelial function of COVID-19 patients
Gabriele Guglielmetti, Marco Quaglia, Pier Paolo Sainaghi, Luigi Mario Castello, Rosanna Vaschetto, Mario Pirisi, Francesco Della Corte, Gian Carlo Avanzi, Piero Stratta, Vincenzo Cantaluppi Critical Care (2020-06-19) https://doi.org/gg35w7
DOI: 10.1186/s13054-020-03060-9 · PMID: 32560665 · PMCID: PMC7303575
125. The complement system in COVID-19: friend and foe?
Anuja Java, Anthony J. Apicelli, M. Kathryn Liszewski, Ariella Coler-Reilly, John P. Atkinson, Alfred H. J. Kim, Hrishikesh S. Kulkarni JCI Insight (2020-08-06) https://doi.org/gg4b5b
DOI: 10.1172/jci.insight.140711 · PMID: 32554923 · PMCID: PMC7455060
126. COVID-19, microangiopathy, hemostatic activation, and complement
Wen-Chao Song, Garret A. FitzGerald Journal of Clinical Investigation (2020-06-22) https://doi.org/gg4b5c
DOI: 10.1172/jci140183 · PMID: 32459663 · PMCID: PMC7410042
127. SARS-CoV-2 infection in primary schools in northern France: A retrospective cohort study in an area of high transmission
Arnaud Fontanet, Rebecca Grant, Laura Tondeur, Yoann Madec, Ludivine Grzelak, Isabelle Cailleau, Marie-Noëlle Ungeheuer, Charlotte Renaudat, Sandrine Fernandes Pellerin, Lucie Kuhmel, … Bruno Hoen Cold Spring Harbor Laboratory (2020-06-29) https://doi.org/gg87nn
DOI: 10.1101/2020.06.25.20140178
128. SARS-CoV-2 Infection in Children
Xiaoxia Lu, Liqiong Zhang, Hui Du, Jingjing Zhang, Yuan Y. Li, Jingyu Qu, Wenxin Zhang, Youjie Wang, Shuangshuang Bao, Ying Li, … Gary W. K. Wong New England Journal of Medicine (2020-04-23) https://doi.org/ggpt2q
DOI: 10.1056/nejmc2005073 · PMID: 32187458 · PMCID: PMC7121177
129. Systematic review of COVID‐19 in children shows milder cases and a better prognosis than adults
Jonas F. Ludvigsson Acta Paediatrica (2020-04-14) https://doi.org/ggq8wr
DOI: 10.1111/apa.15270 · PMID: 32202343 · PMCID: PMC7228328
131. Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Infection in Children and Adolescents
Riccardo Castagnoli, Martina Votto, Amelia Licari, Ilaria Brambilla, Raffaele Bruno, Stefano Perlini, Francesca Rovida, Fausto Baldanti, Gian Luigi Marseglia JAMA Pediatrics (2020-09-01) https://doi.org/dswz
DOI: 10.1001/jamapediatrics.2020.1467 · PMID: 32320004
132. Neurologic and Radiographic Findings Associated With COVID-19 Infection in Children
Omar Abdel-Mannan, Michael Eyre, Ulrike Löbel, Alasdair Bamford, Christin Eltze, Biju Hameed, Cheryl Hemingway, Yael Hacohen JAMA Neurology (2020-11-01) https://doi.org/gg339f
DOI: 10.1001/jamaneurol.2020.2687 · PMID: 32609336 · PMCID: PMC7330822
133. Children with Covid-19 in Pediatric Emergency Departments in Italy
Niccolò Parri, Matteo Lenge, Danilo Buonsenso New England Journal of Medicine (2020-07-09) https://doi.org/ggtp6z
DOI: 10.1056/nejmc2007617 · PMID: 32356945 · PMCID: PMC7206930
134. COVID-19 in 7780 pediatric patients: A systematic review
Ansel Hoang, Kevin Chorath, Axel Moreira, Mary Evans, Finn Burmeister-Morton, Fiona Burmeister, Rija Naqvi, Matthew Petershack, Alvaro Moreira EClinicalMedicine (2020-07) https://doi.org/gg4hn2
DOI: 10.1016/j.eclinm.2020.100433 · PMID: 32766542 · PMCID: PMC7318942
135. Multisystem Inflammatory Syndrome in Children During the Coronavirus 2019 Pandemic: A Case Series
Kathleen Chiotos, Hamid Bassiri, Edward M Behrens, Allison M Blatz, Joyce Chang, Caroline Diorio, Julie C Fitzgerald, Alexis Topjian, Audrey R Odom John Journal of the Pediatric Infectious Diseases Society (2020-07) https://doi.org/ggx4pd
DOI: 10.1093/jpids/piaa069 · PMID: 32463092 · PMCID: PMC7313950
136. Clinical Characteristics of 58 Children With a Pediatric Inflammatory Multisystem Syndrome Temporally Associated With SARS-CoV-2
Elizabeth Whittaker, Alasdair Bamford, Julia Kenny, Myrsini Kaforou, Christine E. Jones, Priyen Shah, Padmanabhan Ramnarayan, Alain Fraisse, Owen Miller, Patrick Davies, … for the PIMS-TS Study Group and EUCLIDS and PERFORM Consortia JAMA (2020-07-21) https://doi.org/gg2v75
DOI: 10.1001/jama.2020.10369 · PMID: 32511692 · PMCID: PMC7281356
137. Toxic shock-like syndrome and COVID-19: Multisystem inflammatory syndrome in children (MIS-C)
Andrea G. Greene, Mona Saleh, Eric Roseman, Richard Sinert The American Journal of Emergency Medicine (2020-11) https://doi.org/gg2586
DOI: 10.1016/j.ajem.2020.05.117 · PMID: 32532619 · PMCID: PMC7274960
138. Multisystem inflammatory syndrome in children and COVID-19 are distinct presentations of SARS–CoV-2
Caroline Diorio, Sarah E. Henrickson, Laura A. Vella, Kevin O. McNerney, Julie Chase, Chakkapong Burudpakdee, Jessica H. Lee, Cristina Jasen, Fran Balamuth, David M. Barrett, … Hamid Bassiri Journal of Clinical Investigation (2020-10-05) https://doi.org/gg7mz2
DOI: 10.1172/jci140970 · PMID: 32730233 · PMCID: PMC7598044
139. The Immunology of Multisystem Inflammatory Syndrome in Children with COVID-19
Camila Rosat Consiglio, Nicola Cotugno, Fabian Sardh, Christian Pou, Donato Amodio, Lucie Rodriguez, Ziyang Tan, Sonia Zicari, Alessandra Ruggiero, Giuseppe Rubens Pascucci, … Petter Brodin Cell (2020-11) https://doi.org/d8fh
DOI: 10.1016/j.cell.2020.09.016 · PMID: 32966765 · PMCID: PMC7474869
140. Acute Heart Failure in Multisystem Inflammatory Syndrome in Children in the Context of Global SARS-CoV-2 Pandemic
Zahra Belhadjer, Mathilde Méot, Fanny Bajolle, Diala Khraiche, Antoine Legendre, Samya Abakka, Johanne Auriau, Marion Grimaud, Mehdi Oualha, Maurice Beghetti, … Damien Bonnet Circulation (2020-08-04) https://doi.org/ggwkv6
DOI: 10.1161/circulationaha.120.048360 · PMID: 32418446
141. An adult with Kawasaki-like multisystem inflammatory syndrome associated with COVID-19
Sheila Shaigany, Marlis Gnirke, Allison Guttmann, Hong Chong, Shane Meehan, Vanessa Raabe, Eddie Louie, Bruce Solitar, Alisa Femia The Lancet (2020-07) https://doi.org/gg4sd6
DOI: 10.1016/s0140-6736(20)31526-9 · PMID: 32659211 · PMCID: PMC7351414
142. COVID-19 associated Kawasaki-like multisystem inflammatory disease in an adult
Sabrina Sokolovsky, Parita Soni, Taryn Hoffman, Philip Kahn, Joshua Scheers-Masters The American Journal of Emergency Medicine (2021-01) https://doi.org/gg5tf4
DOI: 10.1016/j.ajem.2020.06.053 · PMID: 32631771 · PMCID: PMC7315983
143. Characteristics and Outcomes of US Children and Adolescents With Multisystem Inflammatory Syndrome in Children (MIS-C) Compared With Severe Acute COVID-19
Leora R. Feldstein, Mark W. Tenforde, Kevin G. Friedman, Margaret Newhams, Erica Billig Rose, Heda Dapul, Vijaya L. Soma, Aline B. Maddux, Peter M. Mourani, Cindy Bowens, … Overcoming COVID-19 Investigators JAMA (2021-03-16) https://doi.org/gh599q
DOI: 10.1001/jama.2021.2091 · PMID: 33625505 · PMCID: PMC7905703
144. Molecular biology of the cell
Bruce Alberts (editor) Garland Science (2002)
ISBN: 9780815332183
145. Vander’s human physiology: the mechanisms of body function
Eric P Widmaier, Hershel Raff, Kevin T Strang McGraw-Hill Higher Education (2008)
ISBN: 9780071283663
148. Into the Eye of the Cytokine Storm
J. R. Tisoncik, M. J. Korth, C. P. Simmons, J. Farrar, T. R. Martin, M. G. Katze Microbiology and Molecular Biology Reviews (2012-03-05) https://doi.org/f4n9h2
DOI: 10.1128/mmbr.05015-11 · PMID: 22390970 · PMCID: PMC3294426
150. Dynamic balance of pro- and anti-inflammatory signals controls disease and limits pathology
Joseph M. Cicchese, Stephanie Evans, Caitlin Hult, Louis R. Joslyn, Timothy Wessler, Jess A. Millar, Simeone Marino, Nicholas A. Cilfone, Joshua T. Mattila, Jennifer J. Linderman, Denise E. Kirschner Immunological Reviews (2018-09) https://doi.org/gd4g4p
DOI: 10.1111/imr.12671 · PMID: 30129209 · PMCID: PMC6292442
151. Cytokine Dysregulation, Inflammation and Well-Being
Ilia J. Elenkov, Domenic G. Iezzoni, Adrian Daly, Alan G. Harris, George P. Chrousos Neuroimmunomodulation (2005) https://doi.org/bsn7kn
DOI: 10.1159/000087104 · PMID: 16166805
152. Chronic inflammation in the etiology of disease across the life span
David Furman, Judith Campisi, Eric Verdin, Pedro Carrera-Bastos, Sasha Targ, Claudio Franceschi, Luigi Ferrucci, Derek W. Gilroy, Alessio Fasano, Gary W. Miller, … George M. Slavich Nature Medicine (2019-12-05) https://doi.org/gghdcf
DOI: 10.1038/s41591-019-0675-0 · PMID: 31806905 · PMCID: PMC7147972
153. Inpatient care for septicemia or sepsis: a challenge for patients and hospitals.
Margaret Jean Hall, Sonja N Williams, Carol J DeFrances, Aleksandr Golosinskiy NCHS data brief (2011-06) https://www.ncbi.nlm.nih.gov/pubmed/22142805
PMID: 22142805
154. Cytokine Balance in the Lungs of Patients with Acute Respiratory Distress Syndrome
WILLIAM Y. PARK, RICHARD B. GOODMAN, KENNETH P. STEINBERG, JOHN T. RUZINSKI, FRANK RADELLA, DAVID R. PARK, JEROME PUGIN, SHAWN J. SKERRETT, LEONARD D. HUDSON, THOMAS R. MARTIN American Journal of Respiratory and Critical Care Medicine (2001-11-15) https://doi.org/ggqfq7
DOI: 10.1164/ajrccm.164.10.2104013 · PMID: 11734443
155. Cytokine release syndrome
Alexander Shimabukuro-Vornhagen, Philipp Gödel, Marion Subklewe, Hans Joachim Stemmler, Hans Anton Schlößer, Max Schlaak, Matthias Kochanek, Boris Böll, Michael S. von Bergwelt-Baildon Journal for ImmunoTherapy of Cancer (2018-06-15) https://doi.org/ghbncj
DOI: 10.1186/s40425-018-0343-9 · PMID: 29907163 · PMCID: PMC6003181
157. The pro- and anti-inflammatory properties of the cytokine interleukin-6
Jürgen Scheller, Athena Chalaris, Dirk Schmidt-Arras, Stefan Rose-John Biochimica et Biophysica Acta (BBA) - Molecular Cell Research (2011-05) https://doi.org/cvn4nr
DOI: 10.1016/j.bbamcr.2011.01.034 · PMID: 21296109
158. The Role of Interleukin 6 During Viral Infections
Lauro Velazquez-Salinas, Antonio Verdugo-Rodriguez, Luis L. Rodriguez, Manuel V. Borca Frontiers in Microbiology (2019-05-10) https://doi.org/ghbnck
DOI: 10.3389/fmicb.2019.01057 · PMID: 31134045 · PMCID: PMC6524401
159. Expression of elevated levels of pro-inflammatory cytokines in SARS-CoV-infected ACE2 + cells in SARS patients: relation to the acute lung injury and pathogenesis of SARS
L He, Y Ding, Q Zhang, X Che, Y He, H Shen, H Wang, Z Li, L Zhao, J Geng, … S Jiang The Journal of Pathology (2006-11) https://doi.org/bwb8ns
DOI: 10.1002/path.2067 · PMID: 17031779
160. Up-regulation of IL-6 and TNF-α induced by SARS-coronavirus spike protein in murine macrophages via NF-κB pathway
Wei Wang, Linbai Ye, Li Ye, Baozong Li, Bo Gao, Yingchun Zeng, Lingbao Kong, Xiaonan Fang, Hong Zheng, Zhenghui Wu, Yinglong She Virus Research (2007-09) https://doi.org/bm7m55
DOI: 10.1016/j.virusres.2007.02.007 · PMID: 17532082 · PMCID: PMC7114322
161. COVID-19: consider cytokine storm syndromes and immunosuppression
Puja Mehta, Daniel F McAuley, Michael Brown, Emilie Sanchez, Rachel S Tattersall, Jessica J Manson The Lancet (2020-03) https://doi.org/ggnzmc
DOI: 10.1016/s0140-6736(20)30628-0
164. Can we use interleukin-6 (IL-6) blockade for coronavirus disease 2019 (COVID-19)-induced cytokine release syndrome (CRS)?
Bingwen Liu, Min Li, Zhiguang Zhou, Xuan Guan, Yufei Xiang Journal of Autoimmunity (2020-07) https://doi.org/ggr79c
DOI: 10.1016/j.jaut.2020.102452 · PMID: 32291137 · PMCID: PMC7151347
165. A systems approach to infectious disease
Manon Eckhardt, Judd F. Hultquist, Robyn M. Kaake, Ruth Hüttenhain, Nevan J. Krogan Nature Reviews Genetics (2020-02-14) https://doi.org/ggnv63
DOI: 10.1038/s41576-020-0212-5 · PMID: 32060427
166. Differential expression of serum/plasma proteins in various infectious diseases: Specific or nonspecific signatures
Sandipan Ray, Sandip K. Patel, Vipin Kumar, Jagruti Damahe, Sanjeeva Srivastava PROTEOMICS - Clinical Applications (2014-02) https://doi.org/f2px3h
DOI: 10.1002/prca.201300074 · PMID: 24293340
167. Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19
Daniel Blanco-Melo, Benjamin E. Nilsson-Payant, Wen-Chun Liu, Skyler Uhl, Daisy Hoagland, Rasmus Møller, Tristan X. Jordan, Kohei Oishi, Maryline Panis, David Sachs, … Benjamin R. tenOever Cell (2020-05) https://doi.org/ggw5tq
DOI: 10.1016/j.cell.2020.04.026 · PMID: 32416070 · PMCID: PMC7227586
168. SARS-CoV-2 ORF3b Is a Potent Interferon Antagonist Whose Activity Is Increased by a Naturally Occurring Elongation Variant
Yoriyuki Konno, Izumi Kimura, Keiya Uriu, Masaya Fukushi, Takashi Irie, Yoshio Koyanagi, Daniel Sauter, Robert J. Gifford, So Nakagawa, Kei Sato Cell Reports (2020-09) https://doi.org/ghvf8j
DOI: 10.1016/j.celrep.2020.108185 · PMID: 32941788 · PMCID: PMC7473339
169. Bulk and single-cell gene expression profiling of SARS-CoV-2 infected human cell lines identifies molecular targets for therapeutic intervention
Wyler Emanuel, Mösbauer Kirstin, Franke Vedran, Diag Asija, Gottula Lina Theresa, Arsie Roberto, Klironomos Filippos, Koppstein David, Ayoub Salah, Buccitelli Christopher, … Landthaler Markus Cold Spring Harbor Laboratory (2020-05-05) https://doi.org/ggxd2g
DOI: 10.1101/2020.05.05.079194
170. Isolation and characterization of SARS-CoV-2 from the first US COVID-19 patient
Jennifer Harcourt, Azaibi Tamin, Xiaoyan Lu, Shifaq Kamili, Senthil Kumar. Sakthivel, Janna Murray, Krista Queen, Ying Tao, Clinton R. Paden, Jing Zhang, … Natalie J. Thornburg Cold Spring Harbor Laboratory (2020-03-07) https://doi.org/gg2fkm
DOI: 10.1101/2020.03.02.972935 · PMID: 32511316 · PMCID: PMC7239045
171. Proteomics of SARS-CoV-2-infected host cells reveals therapy targets
Denisa Bojkova, Kevin Klann, Benjamin Koch, Marek Widera, David Krause, Sandra Ciesek, Jindrich Cinatl, Christian Münch Nature (2020-05-14) https://doi.org/dw7s
DOI: 10.1038/s41586-020-2332-7 · PMID: 32408336
172. Potent human neutralizing antibodies elicited by SARS-CoV-2 infection
Bin Ju, Qi Zhang, Xiangyang Ge, Ruoke Wang, Jiazhen Yu, Sisi Shan, Bing Zhou, Shuo Song, Xian Tang, Jinfang Yu, … Linqi Zhang Cold Spring Harbor Laboratory (2020-03-26) https://doi.org/ggp7t4
DOI: 10.1101/2020.03.21.990770
173. Plasma proteome of severe acute respiratory syndrome analyzed by two-dimensional gel electrophoresis and mass spectrometry
J.-H. Chen, Y.-W. Chang, C.-W. Yao, T.-S. Chiueh, S.-C. Huang, K.-Y. Chien, A. Chen, F.-Y. Chang, C.-H. Wong, Y.-J. Chen Proceedings of the National Academy of Sciences (2004-11-30) https://doi.org/dtv8sx
DOI: 10.1073/pnas.0407992101 · PMID: 15572443 · PMCID: PMC535397
174. Analysis of multimerization of the SARS coronavirus nucleocapsid protein
Runtao He, Frederick Dobie, Melissa Ballantine, Andrew Leeson, Yan Li, Nathalie Bastien, Todd Cutts, Anton Andonov, Jingxin Cao, Timothy F. Booth, … Xuguang Li Biochemical and Biophysical Research Communications (2004-04) https://doi.org/dbfwr9
DOI: 10.1016/j.bbrc.2004.02.074 · PMID: 15020242 · PMCID: PMC7111152
177. The Immune Epitope Database (IEDB): 2018 update
Randi Vita, Swapnil Mahajan, James A Overton, Sandeep Kumar Dhanda, Sheridan Martini, Jason R Cantrell, Daniel K Wheeler, Alessandro Sette, Bjoern Peters Nucleic Acids Research (2019-01-08) https://doi.org/gfhz6n
DOI: 10.1093/nar/gky1006 · PMID: 30357391 · PMCID: PMC6324067
178. ViPR: an open bioinformatics database and analysis resource for virology research
Brett E. Pickett, Eva L. Sadat, Yun Zhang, Jyothi M. Noronha, R. Burke Squires, Victoria Hunt, Mengya Liu, Sanjeev Kumar, Sam Zaremba, Zhiping Gu, … Richard H. Scheuermann Nucleic Acids Research (2012-01) https://doi.org/c3tds5
DOI: 10.1093/nar/gkr859 · PMID: 22006842 · PMCID: PMC3245011
179. A SARS-CoV-2-Human Protein-Protein Interaction Map Reveals Drug Targets and Potential Drug-Repurposing
David E. Gordon, Gwendolyn M. Jang, Mehdi Bouhaddou, Jiewei Xu, Kirsten Obernier, Matthew J. O’Meara, Jeffrey Z. Guo, Danielle L. Swaney, Tia A. Tummino, Ruth Hüttenhain, … Nevan J. Krogan Cold Spring Harbor Laboratory (2020-03-22) https://doi.org/ggpptg
DOI: 10.1101/2020.03.22.002386 · PMID: 32511329
180. Protein Palmitoylation and Its Role in Bacterial and Viral Infections
Justyna Sobocińska, Paula Roszczenko-Jasińska, Anna Ciesielska, Katarzyna Kwiatkowska Frontiers in Immunology (2018-01-19) https://doi.org/gcxpp2
DOI: 10.3389/fimmu.2017.02003 · PMID: 29403483 · PMCID: PMC5780409
183. The severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) envelope (E) protein harbors a conserved BH3-like sequence
Vincent Navratil, Loïc Lionnard, Sonia Longhi, J. Marie Hardwick, Christophe Combet, Abdel Aouacheria Cold Spring Harbor Laboratory (2020-06-09) https://doi.org/ggrp43
DOI: 10.1101/2020.04.09.033522
185. Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2
Renhong Yan, Yuanyuan Zhang, Yaning Li, Lu Xia, Yingying Guo, Qiang Zhou Science (2020-03-27) https://doi.org/ggpxc8
DOI: 10.1126/science.abb2762 · PMID: 32132184 · PMCID: PMC7164635
186. Structural basis of receptor recognition by SARS-CoV-2
Jian Shang, Gang Ye, Ke Shi, Yushun Wan, Chuming Luo, Hideki Aihara, Qibin Geng, Ashley Auerbach, Fang Li Nature (2020-03-30) https://doi.org/ggqspv
DOI: 10.1038/s41586-020-2179-y · PMID: 32225175
187. Crystal structure of the 2019-nCoV spike receptor-binding domain bound with the ACE2 receptor
Jun Lan, Jiwan Ge, Jinfang Yu, Sisi Shan, Huan Zhou, Shilong Fan, Qi Zhang, Xuanling Shi, Qisheng Wang, Linqi Zhang, Xinquan Wang Cold Spring Harbor Laboratory (2020-02-20) https://doi.org/ggqzp5
DOI: 10.1101/2020.02.19.956235
188. Structural and Functional Basis of SARS-CoV-2 Entry by Using Human ACE2
Qihui Wang, Yanfang Zhang, Lili Wu, Sheng Niu, Chunli Song, Zengyuan Zhang, Guangwen Lu, Chengpeng Qiao, Yu Hu, Kwok-Yung Yuen, … Jianxun Qi Cell (2020-05) https://doi.org/ggr2cz
DOI: 10.1016/j.cell.2020.03.045 · PMID: 32275855 · PMCID: PMC7144619
189. Receptor Recognition by the Novel Coronavirus from Wuhan: an Analysis Based on Decade-Long Structural Studies of SARS Coronavirus
Yushun Wan, Jian Shang, Rachel Graham, Ralph S. Baric, Fang Li Journal of Virology (2020-03-17) https://doi.org/ggjvwn
DOI: 10.1128/jvi.00127-20 · PMID: 31996437 · PMCID: PMC7081895
190. Structure of the Hemagglutinin Precursor Cleavage Site, a Determinant of Influenza Pathogenicity and the Origin of the Labile Conformation
Jue Chen, Kon Ho Lee, David A Steinhauer, David J Stevens, John J Skehel, Don C Wiley Cell (1998-10) https://doi.org/bvgh5b
DOI: 10.1016/s0092-8674(00)81771-7
192. Tracking Changes in SARS-CoV-2 Spike: Evidence that D614G Increases Infectivity of the COVID-19 Virus
Bette Korber, Will M. Fischer, Sandrasegaram Gnanakaran, Hyejin Yoon, James Theiler, Werner Abfalterer, Nick Hengartner, Elena E. Giorgi, Tanmoy Bhattacharya, Brian Foley, … Matthew D. Wyles Cell (2020-08) https://doi.org/gg3wqn
DOI: 10.1016/j.cell.2020.06.043 · PMID: 32697968 · PMCID: PMC7332439
193. Structural and Functional Analysis of the D614G SARS-CoV-2 Spike Protein Variant
Leonid Yurkovetskiy, Xue Wang, Kristen E. Pascal, Christopher Tomkins-Tinch, Thomas P. Nyalile, Yetao Wang, Alina Baum, William E. Diehl, Ann Dauphin, Claudia Carbone, … Jeremy Luban Cell (2020-10) https://doi.org/ghkt47
DOI: 10.1016/j.cell.2020.09.032 · PMID: 32991842 · PMCID: PMC7492024
196. SARS-CoV-2 Variant Under Investigation 202012/01 has more than twofold replicative advantage
Frederic Grabowski, Grzegorz Preibisch, Marek Kochańczyk, Tomasz Lipniacki Cold Spring Harbor Laboratory (2021-01-04) https://doi.org/ghtzw3
DOI: 10.1101/2020.12.28.20248906
198. SARS-CoV-2 spike D614G change enhances replication and transmission
Bin Zhou, Tran Thi Nhu Thao, Donata Hoffmann, Adriano Taddeo, Nadine Ebert, Fabien Labroussaa, Anne Pohlmann, Jacqueline King, Silvio Steiner, Jenna N. Kelly, … Martin Beer Nature (2021-02-26) https://doi.org/gh56mp
DOI: 10.1038/s41586-021-03361-1 · PMID: 33636719
199. Controversy around airborne versus droplet transmission of respiratory viruses
Eunice Y. C. Shiu, Nancy H. L. Leung, Benjamin J. Cowling Current Opinion in Infectious Diseases (2019-08) https://doi.org/ggbwdb
DOI: 10.1097/qco.0000000000000563
200. Viral infections acquired indoors through airborne, droplet or contact transmission.
Giuseppina La Rosa, Marta Fratini, Simonetta Della Libera, Marcello Iaconelli, Michele Muscillo Annali dell’Istituto superiore di sanita (2013) https://www.ncbi.nlm.nih.gov/pubmed/23771256
DOI: 10.4415/ann_13_02_03 · PMID: 23771256
203. Assessing the Dynamics and Control of Droplet- and Aerosol-Transmitted Influenza Using an Indoor Positioning System
Timo Smieszek, Gianrocco Lazzari, Marcel Salathé Scientific Reports (2019-02-18) https://doi.org/ggnqbc
DOI: 10.1038/s41598-019-38825-y · PMID: 30778136 · PMCID: PMC6379436
204. Influenza A virus transmission via respiratory aerosols or droplets as it relates to pandemic potential
Mathilde Richard, Ron A. M. Fouchier FEMS Microbiology Reviews (2016-01) https://doi.org/f8cp4h
DOI: 10.1093/femsre/fuv039 · PMID: 26385895 · PMCID: PMC5006288
207. Stability of Middle East respiratory syndrome coronavirus (MERS-CoV) under different environmental conditions
N van Doremalen, T Bushmaker, VJ Munster Eurosurveillance (2013-09-19) https://doi.org/ggnnjt
DOI: 10.2807/1560-7917.es2013.18.38.20590 · PMID: 24084338
211. Aerosol and Surface Stability of SARS-CoV-2 as Compared with SARS-CoV-1
Neeltje van Doremalen, Trenton Bushmaker, Dylan H. Morris, Myndi G. Holbrook, Amandine Gamble, Brandi N. Williamson, Azaibi Tamin, Jennifer L. Harcourt, Natalie J. Thornburg, Susan I. Gerber, … Vincent J. Munster New England Journal of Medicine (2020-04-16) https://doi.org/ggn88w
DOI: 10.1056/nejmc2004973 · PMID: 32182409 · PMCID: PMC7121658
212. It Is Time to Address Airborne Transmission of Coronavirus Disease 2019 (COVID-19)
Lidia Morawska, Donald K Milton Clinical Infectious Diseases (2020-07-06) https://doi.org/gg34zn
DOI: 10.1093/cid/ciaa939 · PMID: 32628269 · PMCID: PMC7454469
213. Aerodynamic analysis of SARS-CoV-2 in two Wuhan hospitals
Yuan Liu, Zhi Ning, Yu Chen, Ming Guo, Yingle Liu, Nirmal Kumar Gali, Li Sun, Yusen Duan, Jing Cai, Dane Westerdahl, … Ke Lan Nature (2020-04-27) https://doi.org/ggtgng
DOI: 10.1038/s41586-020-2271-3 · PMID: 32340022
216. Clinical characteristics of 24 asymptomatic infections with COVID-19 screened among close contacts in Nanjing, China
Zhiliang Hu, Ci Song, Chuanjun Xu, Guangfu Jin, Yaling Chen, Xin Xu, Hongxia Ma, Wei Chen, Yuan Lin, Yishan Zheng, … Hongbing Shen Science China Life Sciences (2020-03-04) https://doi.org/dqbn
DOI: 10.1007/s11427-020-1661-4 · PMID: 32146694 · PMCID: PMC7088568
217. Evidence for transmission of COVID-19 prior to symptom onset
Lauren C Tindale, Jessica E Stockdale, Michelle Coombe, Emma S Garlock, Wing Yin Venus Lau, Manu Saraswat, Louxin Zhang, Dongxuan Chen, Jacco Wallinga, Caroline Colijn eLife (2020-06-22) https://doi.org/gg6dtw
DOI: 10.7554/elife.57149 · PMID: 32568070 · PMCID: PMC7386904
218. Time Kinetics of Viral Clearance and Resolution of Symptoms in Novel Coronavirus Infection
De Chang, Guoxin Mo, Xin Yuan, Yi Tao, Xiaohua Peng, Fu-Sheng Wang, Lixin Xie, Lokesh Sharma, Charles S. Dela Cruz, Enqiang Qin American Journal of Respiratory and Critical Care Medicine (2020-05-01) https://doi.org/ggq8xs
DOI: 10.1164/rccm.202003-0524le · PMID: 32200654 · PMCID: PMC7193851
219. Temporal dynamics in viral shedding and transmissibility of COVID-19
Xi He, Eric H. Y. Lau, Peng Wu, Xilong Deng, Jian Wang, Xinxin Hao, Yiu Chung Lau, Jessica Y. Wong, Yujuan Guan, Xinghua Tan, … Gabriel M. Leung Nature Medicine (2020-04-15) https://doi.org/ggr99q
DOI: 10.1038/s41591-020-0869-5 · PMID: 32296168
221. Virological assessment of hospitalized patients with COVID-2019
Roman Wölfel, Victor M. Corman, Wolfgang Guggemos, Michael Seilmaier, Sabine Zange, Marcel A. Müller, Daniela Niemeyer, Terry C. Jones, Patrick Vollmar, Camilla Rothe, … Clemens Wendtner Nature (2020-04-01) https://doi.org/ggqrv7
DOI: 10.1038/s41586-020-2196-x · PMID: 32235945
222. Clinical predictors and timing of cessation of viral RNA shedding in patients with COVID-19
Cristina Corsini Campioli, Edison Cano Cevallos, Mariam Assi, Robin Patel, Matthew J. Binnicker, John C. O’Horo Journal of Clinical Virology (2020-09) https://doi.org/gg7m96
DOI: 10.1016/j.jcv.2020.104577 · PMID: 32777762 · PMCID: PMC7405830
223. Presymptomatic SARS-CoV-2 Infections and Transmission in a Skilled Nursing Facility
Melissa M. Arons, Kelly M. Hatfield, Sujan C. Reddy, Anne Kimball, Allison James, Jesica R. Jacobs, Joanne Taylor, Kevin Spicer, Ana C. Bardossy, Lisa P. Oakley, … John A. Jernigan New England Journal of Medicine (2020-05-28) https://doi.org/ggszfg
DOI: 10.1056/nejmoa2008457 · PMID: 32329971 · PMCID: PMC7200056
224. Prevalence of SARS-CoV-2 Infection in Residents of a Large Homeless Shelter in Boston
Travis P. Baggett, Harrison Keyes, Nora Sporn, Jessie M. Gaeta JAMA (2020-06-02) https://doi.org/ggtsh3
DOI: 10.1001/jama.2020.6887 · PMID: 32338732 · PMCID: PMC7186911
226. Transmission of COVID-19 in the terminal stages of the incubation period: A familial cluster
Peng Li, Ji-Bo Fu, Ke-Feng Li, Jie-Nan Liu, Hong-Ling Wang, Lei-Jie Liu, Yan Chen, Yong-Li Zhang, She-Lan Liu, An Tang, … Jian-Bo Yan International Journal of Infectious Diseases (2020-07) https://doi.org/ggq844
DOI: 10.1016/j.ijid.2020.03.027 · PMID: 32194239 · PMCID: PMC7264481
227. A Cohort of SARS-CoV-2 Infected Asymptomatic and Pre-Symptomatic Contacts from COVID-19 Contact Tracing in Hubei Province, China: Short-Term Outcomes
Peng Zhang, Fei Tian, Yuan Wan, Jing Cai, Zhengmin Qian, Ran Wu, Yunquan Zhang, Shiyu Zhang, Huan Li, Mingyan Li, … Hualiang Lin SSRN Electronic Journal (2020) https://doi.org/ghf3n2
DOI: 10.2139/ssrn.3678556
228. Estimating the asymptomatic proportion of coronavirus disease 2019 (COVID-19) cases on board the Diamond Princess cruise ship, Yokohama, Japan, 2020
Kenji Mizumoto, Katsushi Kagaya, Alexander Zarebski, Gerardo Chowell Eurosurveillance (2020-03-12) https://doi.org/ggn4bd
DOI: 10.2807/1560-7917.es.2020.25.10.2000180 · PMID: 32183930 · PMCID: PMC7078829
229. Estimated prevalence and viral transmissibility in subjects with asymptomatic SARS-CoV-2 infections in Wuhan, China
Kang Zhang, Weiwei Tong, Xinghuan Wang, Johnson Yiu-Nam Lau Precision Clinical Medicine (2020-12) https://doi.org/ghjmks
DOI: 10.1093/pcmedi/pbaa032 · PMCID: PMC7499683
230. Investigation of SARS-CoV-2 outbreaks in six care homes in London, April 2020
Shamez N Ladhani, J.Yimmy Chow, Roshni Janarthanan, Jonathan Fok, Emma Crawley-Boevey, Amoolya Vusirikala, Elena Fernandez, Marina Sanchez Perez, Suzanne Tang, Kate Dun-Campbell, … Maria Zambon EClinicalMedicine (2020-09) https://doi.org/ghbj9v
DOI: 10.1016/j.eclinm.2020.100533 · PMID: 32923993 · PMCID: PMC7480335
232. Suppression of a SARS-CoV-2 outbreak in the Italian municipality of Vo’
Enrico Lavezzo, Elisa Franchin, Constanze Ciavarella, Gina Cuomo-Dannenburg, Luisa Barzon, Claudia Del Vecchio, Lucia Rossi, Riccardo Manganelli, Arianna Loregian, Nicolò Navarin, … Imperial College COVID-19 Response Team Nature (2020-06-30) https://doi.org/gg3w87
DOI: 10.1038/s41586-020-2488-1 · PMID: 32604404
233. A systematic review and meta-analysis of published research data on COVID-19 infection fatality rates
Gideon Meyerowitz-Katz, Lea Merone International Journal of Infectious Diseases (2020-12) https://doi.org/ghgjpw
DOI: 10.1016/j.ijid.2020.09.1464 · PMID: 33007452 · PMCID: PMC7524446
236. Repeated cross-sectional sero-monitoring of SARS-CoV-2 in New York City
Daniel Stadlbauer, Jessica Tan, Kaijun Jiang, Matthew M. Hernandez, Shelcie Fabre, Fatima Amanat, Catherine Teo, Guha Asthagiri Arunkumar, Meagan McMahon, Christina Capuano, … Florian Krammer Nature (2020-11-03) https://doi.org/ghhtq9
DOI: 10.1038/s41586-020-2912-6 · PMID: 33142304
239. Modeling infectious diseases in humans and animals
Matthew James Keeling, Pejman Rohani Princeton University Press (2008)
ISBN: 9780691116174
240. A contribution to the mathematical theory of epidemics
Proceedings of the Royal Society of London. Series A, Containing Papers of a Mathematical and Physical Character
(1997-01) https://doi.org/fwx2qw
DOI: 10.1098/rspa.1927.0118
243. Theoretical ecology: principles and applications
Robert M. May, Angela R. McLean (editors) Oxford University Press (2007)
ISBN: 9780199209989
244. Nowcasting and forecasting the potential domestic and international spread of the 2019-nCoV outbreak originating in Wuhan, China: a modelling study
Joseph T Wu, Kathy Leung, Gabriel M Leung The Lancet (2020-02) https://doi.org/ggjvr7
DOI: 10.1016/s0140-6736(20)30260-9
245. The reproductive number of COVID-19 is higher compared to SARS coronavirus
Ying Liu, Albert A Gayle, Annelies Wilder-Smith, Joacim Rocklöv Journal of Travel Medicine (2020-03) https://doi.org/ggnntv
DOI: 10.1093/jtm/taaa021 · PMID: 32052846 · PMCID: PMC7074654
246. Substantial undocumented infection facilitates the rapid dissemination of novel coronavirus (SARS-CoV-2)
Ruiyun Li, Sen Pei, Bin Chen, Yimeng Song, Tao Zhang, Wan Yang, Jeffrey Shaman Science (2020-05-01) https://doi.org/ggn6c2
DOI: 10.1126/science.abb3221 · PMID: 32179701
247. Epidemiological parameters of coronavirus disease 2019: a pooled analysis of publicly reported individual data of 1155 cases from seven countries
Shujuan Ma, Jiayue Zhang, Minyan Zeng, Qingping Yun, Wei Guo, Yixiang Zheng, Shi Zhao, Maggie H. Wang, Zuyao Yang Cold Spring Harbor Laboratory (2020-03-24) https://doi.org/ggqhzz
DOI: 10.1101/2020.03.21.20040329
248. Early Transmissibility Assessment of a Novel Coronavirus in Wuhan, China
Maimuna Majumder, Kenneth D. Mandl SSRN Electronic Journal (2020) https://doi.org/ggqhz3
DOI: 10.2139/ssrn.3524675 · PMID: 32714102
249. Time-varying transmission dynamics of Novel Coronavirus Pneumonia in China
Tao Liu, Jianxiong Hu, Jianpeng Xiao, Guanhao He, Min Kang, Zuhua Rong, Lifeng Lin, Haojie Zhong, Qiong Huang, Aiping Deng, … Wenjun Ma Cold Spring Harbor Laboratory (2020-02-13) https://doi.org/dkx9
DOI: 10.1101/2020.01.25.919787
250. Estimation of the reproductive number of novel coronavirus (COVID-19) and the probable outbreak size on the Diamond Princess cruise ship: A data-driven analysis
Sheng Zhang, MengYuan Diao, Wenbo Yu, Lei Pei, Zhaofen Lin, Dechang Chen International Journal of Infectious Diseases (2020-04) https://doi.org/ggpx56
DOI: 10.1016/j.ijid.2020.02.033 · PMID: 32097725 · PMCID: PMC7110591
251. Estimation of the Transmission Risk of the 2019-nCoV and Its Implication for Public Health Interventions
Biao Tang, Xia Wang, Qian Li, Nicola Luigi Bragazzi, Sanyi Tang, Yanni Xiao, Jianhong Wu Journal of Clinical Medicine (2020-02-07) https://doi.org/ggmkf4
DOI: 10.3390/jcm9020462 · PMID: 32046137 · PMCID: PMC7074281
253. Modelling the epidemic trend of the 2019 novel coronavirus outbreak in China
Mingwang Shen, Zhihang Peng, Yanni Xiao, Lei Zhang Cold Spring Harbor Laboratory (2020-01-25) https://doi.org/ggqhzw
DOI: 10.1101/2020.01.23.916726
254. Novel coronavirus 2019-nCoV: early estimation of epidemiological parameters and epidemic predictions
Jonathan M. Read, Jessica R. E. Bridgen, Derek A. T. Cummings, Antonia Ho, Chris P. Jewell Cold Spring Harbor Laboratory (2020-01-28) https://doi.org/dkzb
DOI: 10.1101/2020.01.23.20018549
255. Using early data to estimate the actual infection fatality ratio from COVID-19 in France
Lionel Roques, Etienne Klein, Julien Papaïx, Antoine Sar, Samuel Soubeyrand Cold Spring Harbor Laboratory (2020-05-07) https://doi.org/ggqhz2
DOI: 10.1101/2020.03.22.20040915
256. Potential roles of social distancing in mitigating the spread of coronavirus disease 2019 (COVID-19) in South Korea
Sang Woo Park, Kaiyuan Sun, Cécile Viboud, Bryan T Grenfell, Jonathan Dushoff GitHub (2020) https://github.com/parksw3/Korea-analysis/blob/master/v1/korea.pdf
257. Early dynamics of transmission and control of COVID-19: a mathematical modelling study
Adam J Kucharski, Timothy W Russell, Charlie Diamond, Yang Liu, John Edmunds, Sebastian Funk, Rosalind M Eggo, Fiona Sun, Mark Jit, James D Munday, … Stefan Flasche The Lancet Infectious Diseases (2020-05) https://doi.org/ggptcf
DOI: 10.1016/s1473-3099(20)30144-4
258. Estimating the reproduction number of COVID-19 in Iran using epidemic modeling
Ebrahim Sahafizadeh, Samaneh Sartoli Cold Spring Harbor Laboratory (2020-04-23) https://doi.org/ggqhzx
DOI: 10.1101/2020.03.20.20038422
259. Report 13: Estimating the number of infections and the impact of non-pharmaceutical interventions on COVID-19 in 11 European countries
S Flaxman, S Mishra, A Gandy, H Unwin, H Coupland, T Mellan, H Zhu, T Berah, J Eaton, P Perez Guzman, … S Bhatt Imperial College London (2020-03-30) https://doi.org/ggrbmf
DOI: 10.25561/77731
260. Projecting hospital utilization during the COVID-19 outbreaks in the United States
Seyed M. Moghadas, Affan Shoukat, Meagan C. Fitzpatrick, Chad R. Wells, Pratha Sah, Abhishek Pandey, Jeffrey D. Sachs, Zheng Wang, Lauren A. Meyers, Burton H. Singer, Alison P. Galvani Proceedings of the National Academy of Sciences (2020-04-21) https://doi.org/ggq7jc
DOI: 10.1073/pnas.2004064117 · PMID: 32245814 · PMCID: PMC7183199
261. The effect of control strategies to reduce social mixing on outcomes of the COVID-19 epidemic in Wuhan, China: a modelling study
Kiesha Prem, Yang Liu, Timothy W Russell, Adam J Kucharski, Rosalind M Eggo, Nicholas Davies, Mark Jit, Petra Klepac, Stefan Flasche, Samuel Clifford, … Joel Hellewell The Lancet Public Health (2020-05) https://doi.org/ggp3xq
DOI: 10.1016/s2468-2667(20)30073-6 · PMID: 32220655 · PMCID: PMC7158905
262. Spread and dynamics of the COVID-19 epidemic in Italy: Effects of emergency containment measures
Marino Gatto, Enrico Bertuzzo, Lorenzo Mari, Stefano Miccoli, Luca Carraro, Renato Casagrandi, Andrea Rinaldo Proceedings of the National Academy of Sciences (2020-05-12) https://doi.org/ggv4j6
DOI: 10.1073/pnas.2004978117 · PMID: 32327608 · PMCID: PMC7229754
263. Covid-19: Temporal variation in transmission during the COVID-19 outbreak
EpiForecasts and the CMMID Covid working group https://epiforecasts.io/covid/
264. Rt COVID-19
Kevin Systrom, Thomas Vladeck, Mike Krieger https://rt.live/
265. Emerging SARS-CoV-2 mutation hot spots include a novel RNA-dependent-RNA polymerase variant
Maria Pachetti, Bruna Marini, Francesca Benedetti, Fabiola Giudici, Elisabetta Mauro, Paola Storici, Claudio Masciovecchio, Silvia Angeletti, Massimo Ciccozzi, Robert C. Gallo, … Rudy Ippodrino Journal of Translational Medicine (2020-04-22) https://doi.org/ggtzrr
DOI: 10.1186/s12967-020-02344-6 · PMID: 32321524 · PMCID: PMC7174922
266. Emergence of genomic diversity and recurrent mutations in SARS-CoV-2
Lucy van Dorp, Mislav Acman, Damien Richard, Liam P. Shaw, Charlotte E. Ford, Louise Ormond, Christopher J. Owen, Juanita Pang, Cedric C. S. Tan, Florencia A. T. Boshier, … François Balloux Infection, Genetics and Evolution (2020-09) https://doi.org/ggvz4h
DOI: 10.1016/j.meegid.2020.104351 · PMID: 32387564 · PMCID: PMC7199730
268. Coast-to-Coast Spread of SARS-CoV-2 during the Early Epidemic in the United States
Joseph R. Fauver, Mary E. Petrone, Emma B. Hodcroft, Kayoko Shioda, Hanna Y. Ehrlich, Alexander G. Watts, Chantal B. F. Vogels, Anderson F. Brito, Tara Alpert, Anthony Muyombwe, … Nathan D. Grubaugh Cell (2020-05) https://doi.org/gg6r9x
DOI: 10.1016/j.cell.2020.04.021 · PMID: 32386545 · PMCID: PMC7204677
269. Introductions and early spread of SARS-CoV-2 in the New York City area
Ana S. Gonzalez-Reiche, Matthew M. Hernandez, Mitchell J. Sullivan, Brianne Ciferri, Hala Alshammary, Ajay Obla, Shelcie Fabre, Giulio Kleiner, Jose Polanco, Zenab Khan, … Harm van Bakel Science (2020-05-29) https://doi.org/gg5gv7
DOI: 10.1126/science.abc1917 · PMID: 32471856 · PMCID: PMC7259823
270. Spread of SARS-CoV-2 in the Icelandic Population
Daniel F. Gudbjartsson, Agnar Helgason, Hakon Jonsson, Olafur T. Magnusson, Pall Melsted, Gudmundur L. Norddahl, Jona Saemundsdottir, Asgeir Sigurdsson, Patrick Sulem, Arna B. Agustsdottir, … Kari Stefansson New England Journal of Medicine (2020-06-11) https://doi.org/ggr6wx
DOI: 10.1056/nejmoa2006100 · PMID: 32289214 · PMCID: PMC7175425
275. Estimated transmissibility and severity of novel SARS-CoV-2 Variant of Concern 202012/01 in England
Nicholas G. Davies, Rosanna C. Barnard, Christopher I. Jarvis, Adam J. Kucharski, James Munday, Carl A. B. Pearson, Timothy W. Russell, Damien C. Tully, Sam Abbott, Amy Gimma, … CMMID COVID-19 Working Group Cold Spring Harbor Laboratory (2020-12-26) https://doi.org/fp3v
DOI: 10.1101/2020.12.24.20248822
278. Genomic epidemiology identifies emergence and rapid transmission of SARS-CoV-2 B.1.1.7 in the United States
Nicole L. Washington, Karthik Gangavarapu, Mark Zeller, Alexandre Bolze, Elizabeth T. Cirulli, Kelly M. Schiabor Barrett, Brendan B. Larsen, Catelyn Anderson, Simon White, Tyler Cassens, … Kristian G. Andersen Cold Spring Harbor Laboratory (2021-02-07) https://doi.org/gh598v
DOI: 10.1101/2021.02.06.21251159 · PMID: 33564780 · PMCID: PMC7872373
280. Increased mortality in community-tested cases of SARS-CoV-2 lineage B.1.1.7
Nicholas G. Davies, Christopher I. Jarvis, W. John Edmunds, Nicholas P. Jewell, Karla Diaz-Ordaz, Ruth H. Keogh, CMMID COVID-19 Working Group Cold Spring Harbor Laboratory (2021-03-05) https://doi.org/ftkj
DOI: 10.1101/2021.02.01.21250959 · PMID: 33564794 · PMCID: PMC7872389
281. Recurrent emergence and transmission of a SARS-CoV-2 Spike deletion H69/V70
Steven Kemp, William Harvey, Rawlings Datir, Dami Collier, Isabella Ferreira, Bo Meng, Alessandro Carabelii, David L Robertson, Ravindra K Gupta, COVID-19 Genomics UK (COG-UK) consortium Cold Spring Harbor Laboratory (2021-01-13) https://doi.org/ghvq45
DOI: 10.1101/2020.12.14.422555
283. The basis of a more contagious 501Y.V1 variant of SARS-COV-2
Haolin Liu, Qianqian Zhang, Pengcheng Wei, Zhongzhou Chen, Katja Aviszus, John Yang, Walter Downing, Shelley Peterson, Chengyu Jiang, Bo Liang, … Gongyi Zhang Cold Spring Harbor Laboratory (2021-02-02) https://doi.org/gh598t
DOI: 10.1101/2021.02.02.428884 · PMID: 33564771 · PMCID: PMC7872372
284. Epidemiology, transmission dynamics and control of SARS: the 2002–2003 epidemic
Roy M. Anderson, Christophe Fraser, Azra C. Ghani, Christl A. Donnelly, Steven Riley, Neil M. Ferguson, Gabriel M. Leung, T. H. Lam, Anthony J. Hedley Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences (2004-07-29) https://doi.org/c2n646
DOI: 10.1098/rstb.2004.1490 · PMID: 15306395 · PMCID: PMC1693389
286. Human coronavirus circulation in the United States 2014–2017
Marie E. Killerby, Holly M. Biggs, Amber Haynes, Rebecca M. Dahl, Desiree Mustaquim, Susan I. Gerber, John T. Watson Journal of Clinical Virology (2018-04) https://doi.org/gc7sf3
DOI: 10.1016/j.jcv.2018.01.019 · PMID: 29427907 · PMCID: PMC7106380
288. Human (non-severe acute respiratory syndrome) coronavirus infections in hospitalised children in France
Astrid Vabret, Julia Dina, Stéphanie Gouarin, Joëlle Petitjean, Valérie Tripey, Jacques Brouard, François Freymuth Journal of Paediatrics and Child Health (2008-04) https://doi.org/cxt434
DOI: 10.1111/j.1440-1754.2007.01246.x · PMID: 17999671 · PMCID: PMC7166728
290. Case Study: Prolonged Infectious SARS-CoV-2 Shedding from an Asymptomatic Immunocompromised Individual with Cancer
Victoria A. Avanzato, M. Jeremiah Matson, Stephanie N. Seifert, Rhys Pryce, Brandi N. Williamson, Sarah L. Anzick, Kent Barbian, Seth D. Judson, Elizabeth R. Fischer, Craig Martens, … Vincent J. Munster Cell (2020-12) https://doi.org/ghhxkp
DOI: 10.1016/j.cell.2020.10.049 · PMID: 33248470 · PMCID: PMC7640888
291. Persistence and Evolution of SARS-CoV-2 in an Immunocompromised Host
Bina Choi, Manish C. Choudhary, James Regan, Jeffrey A. Sparks, Robert F. Padera, Xueting Qiu, Isaac H. Solomon, Hsiao-Hsuan Kuo, Julie Boucau, Kathryn Bowman, … Jonathan Z. Li New England Journal of Medicine (2020-12-03) https://doi.org/fhv8
DOI: 10.1056/nejmc2031364 · PMID: 33176080 · PMCID: PMC7673303
292. SARS-CoV-2 escape in vitro from a highly neutralizing COVID-19 convalescent plasma
Emanuele Andreano, Giulia Piccini, Danilo Licastro, Lorenzo Casalino, Nicole V. Johnson, Ida Paciello, Simeone Dal Monego, Elisa Pantano, Noemi Manganaro, Alessandro Manenti, … Rino Rappuoli Cold Spring Harbor Laboratory (2020-12-28) https://doi.org/ghs97s
DOI: 10.1101/2020.12.28.424451 · PMID: 33398278 · PMCID: PMC7781313
293. Transmission of SARS-CoV-2 on mink farms between humans and mink and back to humans
Bas B. Oude Munnink, Reina S. Sikkema, David F. Nieuwenhuijse, Robert Jan Molenaar, Emmanuelle Munger, Richard Molenkamp, Arco van der Spek, Paulien Tolsma, Ariene Rietveld, Miranda Brouwer, … Marion P. G. Koopmans Science (2021-01-08) https://doi.org/ghssrq
DOI: 10.1126/science.abe5901 · PMID: 33172935
294. Establishment and lineage dynamics of the SARS-CoV-2 epidemic in the UK
Louis du Plessis, John T. McCrone, Alexander E. Zarebski, Verity Hill, Christopher Ruis, Bernardo Gutierrez, Jayna Raghwani, Jordan Ashworth, Rachel Colquhoun, Thomas R. Connor, … COVID-19 Genomics UK (COG-UK) Consortium† Science (2021-01-08) https://doi.org/ghsbdt
DOI: 10.1126/science.abf2946 · PMID: 33419936
300. Transmission of SARS-CoV-2 Lineage B.1.1.7 in England: Insights from linking epidemiological and genetic data
Erik Volz, Swapnil Mishra, Meera Chand, Jeffrey C. Barrett, Robert Johnson, Lily Geidelberg, Wes R Hinsley, Daniel J Laydon, Gavin Dabrera, Áine O’Toole, … The COVID-19 Genomics UK (COG-UK) consortium Cold Spring Harbor Laboratory (2021-01-04) https://doi.org/ghrqv8
DOI: 10.1101/2020.12.30.20249034
303. S gene dropout patterns in SARS-CoV-2 tests suggest spread of the H69del/V70del mutation in the US
Nicole L. Washington, Simon White, Kelly M. Schiabor Barrett, Elizabeth T. Cirulli, Alexandre Bolze, James T. Lu Cold Spring Harbor Laboratory (2020-12-30) https://doi.org/ghvq46
DOI: 10.1101/2020.12.24.20248814
308. Emergence and rapid spread of a new severe acute respiratory syndrome-related coronavirus 2 (SARS-CoV-2) lineage with multiple spike mutations in South Africa
Houriiyah Tegally, Eduan Wilkinson, Marta Giovanetti, Arash Iranzadeh, Vagner Fonseca, Jennifer Giandhari, Deelan Doolabh, Sureshnee Pillay, Emmanuel James San, Nokukhanya Msomi, … Tulio de Oliveira Cold Spring Harbor Laboratory (2020-12-22) https://doi.org/fqth
DOI: 10.1101/2020.12.21.20248640
314. Emergence of a novel SARS-CoV-2 strain in Southern California, USA
Wenjuan Zhang, Brian D Davis, Stephanie S Chen, Jorge M Sincuir Martinez, Jasmine T Plummer, Eric Vail Cold Spring Harbor Laboratory (2021-01-20) https://doi.org/ghvq48
DOI: 10.1101/2021.01.18.21249786
315. The Impact of Mutations in SARS-CoV-2 Spike on Viral Infectivity and Antigenicity
Qianqian Li, Jiajing Wu, Jianhui Nie, Li Zhang, Huan Hao, Shuo Liu, Chenyan Zhao, Qi Zhang, Huan Liu, Lingling Nie, … Youchun Wang Cell (2020-09) https://doi.org/gg4665
DOI: 10.1016/j.cell.2020.07.012 · PMID: 32730807 · PMCID: PMC7366990
316. Comprehensive mapping of mutations to the SARS-CoV-2 receptor-binding domain that affect recognition by polyclonal human serum antibodies
Allison J. Greaney, Andrea N. Loes, Katharine H. D. Crawford, Tyler N. Starr, Keara D. Malone, Helen Y. Chu, Jesse D. Bloom Cold Spring Harbor Laboratory (2021-01-04) https://doi.org/ghr85d
DOI: 10.1101/2020.12.31.425021
319. Deep Mutational Scanning of SARS-CoV-2 Receptor Binding Domain Reveals Constraints on Folding and ACE2 Binding
Tyler N. Starr, Allison J. Greaney, Sarah K. Hilton, Daniel Ellis, Katharine H. D. Crawford, Adam S. Dingens, Mary Jane Navarro, John E. Bowen, M. Alejandra Tortorici, Alexandra C. Walls, … Jesse D. Bloom Cell (2020-09) https://doi.org/gg72tr
DOI: 10.1016/j.cell.2020.08.012 · PMID: 32841599 · PMCID: PMC7418704
320. Adaptation of SARS-CoV-2 in BALB/c mice for testing vaccine efficacy
Hongjing Gu, Qi Chen, Guan Yang, Lei He, Hang Fan, Yong-Qiang Deng, Yanxiao Wang, Yue Teng, Zhongpeng Zhao, Yujun Cui, … Yusen Zhou Science (2020-07-30) https://doi.org/ghc5mn
DOI: 10.1126/science.abc4730 · PMID: 32732280 · PMCID: PMC7574913
321. Complete Mapping of Mutations to the SARS-CoV-2 Spike Receptor-Binding Domain that Escape Antibody Recognition
Allison J. Greaney, Tyler N. Starr, Pavlo Gilchuk, Seth J. Zost, Elad Binshtein, Andrea N. Loes, Sarah K. Hilton, John Huddleston, Rachel Eguia, Katharine H. D. Crawford, … Jesse D. Bloom Cell Host & Microbe (2021-01) https://doi.org/ghvq3m
DOI: 10.1016/j.chom.2020.11.007 · PMID: 33259788 · PMCID: PMC7676316
322. Recurrent deletions in the SARS-CoV-2 spike glycoprotein drive antibody escape
Kevin R. McCarthy, Linda J. Rennick, Sham Nambulli, Lindsey R. Robinson-McCarthy, William G. Bain, Ghady Haidar, W. Paul Duprex Cold Spring Harbor Laboratory (2021-01-19) https://doi.org/ghvq44
DOI: 10.1101/2020.11.19.389916
326. Prospective mapping of viral mutations that escape antibodies used to treat COVID-19
Tyler N. Starr, Allison J. Greaney, Amin Addetia, William W. Hannon, Manish C. Choudhary, Adam S. Dingens, Jonathan Z. Li, Jesse D. Bloom Science (2021-01-25) https://doi.org/ghvntq
DOI: 10.1126/science.abf9302 · PMID: 33495308
328. mRNA-1273 vaccine induces neutralizing antibodies against spike mutants from global SARS-CoV-2 variants
Kai Wu, Anne P. Werner, Juan I. Moliva, Matthew Koch, Angela Choi, Guillaume B. E. Stewart-Jones, Hamilton Bennett, Seyhan Boyoglu-Barnum, Wei Shi, Barney S. Graham, … Darin K. Edwards Cold Spring Harbor Laboratory (2021-01-25) https://doi.org/fr2g
DOI: 10.1101/2021.01.25.427948 · PMID: 33501442 · PMCID: PMC7836112
329. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine
Fernando P. Polack, Stephen J. Thomas, Nicholas Kitchin, Judith Absalon, Alejandra Gurtman, Stephen Lockhart, John L. Perez, Gonzalo Pérez Marc, Edson D. Moreira, Cristiano Zerbini, … William C. Gruber New England Journal of Medicine (2020-12-31) https://doi.org/ghn625
DOI: 10.1056/nejmoa2034577 · PMID: 33301246 · PMCID: PMC7745181
330. Safety and Immunogenicity of Two RNA-Based Covid-19 Vaccine Candidates
Edward E. Walsh, Robert W. Frenck, Ann R. Falsey, Nicholas Kitchin, Judith Absalon, Alejandra Gurtman, Stephen Lockhart, Kathleen Neuzil, Mark J. Mulligan, Ruth Bailey, … William C. Gruber New England Journal of Medicine (2020-12-17) https://doi.org/ghjktx
DOI: 10.1056/nejmoa2027906 · PMID: 33053279 · PMCID: PMC7583697
331. Neutralization of N501Y mutant SARS-CoV-2 by BNT162b2 vaccine-elicited sera
Xuping Xie, Jing Zou, Camila R. Fontes-Garfias, Hongjie Xia, Kena A. Swanson, Mark Cutler, David Cooper, Vineet D. Menachery, Scott Weaver, Philip R. Dormitzer, Pei-Yong Shi Cold Spring Harbor Laboratory (2021-01-07) https://doi.org/ghvq47
DOI: 10.1101/2021.01.07.425740 · PMID: 33442691 · PMCID: PMC7805448
332. mRNA vaccine-elicited antibodies to SARS-CoV-2 and circulating variants
Zijun Wang, Fabian Schmidt, Yiska Weisblum, Frauke Muecksch, Christopher O. Barnes, Shlomo Finkin, Dennis Schaefer-Babajew, Melissa Cipolla, Christian Gaebler, Jenna A. Lieberman, … Michel C. Nussenzweig Cold Spring Harbor Laboratory (2021-01-30) https://doi.org/frdn
DOI: 10.1101/2021.01.15.426911 · PMID: 33501451 · PMCID: PMC7836122
333. A SARS-CoV-2 vaccine candidate would likely match all currently circulating variants
Bethany Dearlove, Eric Lewitus, Hongjun Bai, Yifan Li, Daniel B. Reeves, M. Gordon Joyce, Paul T. Scott, Mihret F. Amare, Sandhya Vasan, Nelson L. Michael, … Morgane Rolland Proceedings of the National Academy of Sciences (2020-09-22) https://doi.org/fdkz
DOI: 10.1073/pnas.2008281117 · PMID: 32868447 · PMCID: PMC7519301
334. Aggressively find, test, trace and isolate to beat COVID-19
Larissa M. Matukas, Irfan A. Dhalla, Andreas Laupacis Canadian Medical Association Journal (2020-10-05) https://doi.org/gh2jvk
DOI: 10.1503/cmaj.202120 · PMID: 32907821 · PMCID: PMC7546740
335. COVID-19 in New Zealand and the impact of the national response: a descriptive epidemiological study
Sarah Jefferies, Nigel French, Charlotte Gilkison, Giles Graham, Virginia Hope, Jonathan Marshall, Caroline McElnay, Andrea McNeill, Petra Muellner, Shevaun Paine, … Patricia Priest The Lancet Public Health (2020-11) https://doi.org/ftzx
DOI: 10.1016/s2468-2667(20)30225-5 · PMID: 33065023 · PMCID: PMC7553903
336. Potential lessons from the Taiwan and New Zealand health responses to the COVID-19 pandemic
Dr Jennifer Summers, Dr Hao-Yuan Cheng, Professor Hsien-Ho Lin, Dr Lucy Telfar Barnard, Dr Amanda Kvalsvig, Professor Nick Wilson, Professor Michael G Baker The Lancet Regional Health - Western Pacific (2020-11) https://doi.org/ghrzb4
DOI: 10.1016/j.lanwpc.2020.100044 · PMCID: PMC7577184
337. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR
Victor M Corman, Olfert Landt, Marco Kaiser, Richard Molenkamp, Adam Meijer, Daniel KW Chu, Tobias Bleicker, Sebastian Brünink, Julia Schneider, Marie Luisa Schmidt, … Christian Drosten Eurosurveillance (2020-01-23) https://doi.org/ggjs7g
DOI: 10.2807/1560-7917.es.2020.25.3.2000045 · PMID: 31992387 · PMCID: PMC6988269
338. Temporal profiles of viral load in posterior oropharyngeal saliva samples and serum antibody responses during infection by SARS-CoV-2: an observational cohort study
Kelvin Kai-Wang To, Owen Tak-Yin Tsang, Wai-Shing Leung, Anthony Raymond Tam, Tak-Chiu Wu, David Christopher Lung, Cyril Chik-Yan Yip, Jian-Piao Cai, Jacky Man-Chun Chan, Thomas Shiu-Hong Chik, … Kwok-Yung Yuen The Lancet Infectious Diseases (2020-05) https://doi.org/ggp4qx
DOI: 10.1016/s1473-3099(20)30196-1 · PMID: 32213337 · PMCID: PMC7158907
340. Library preparation for next generation sequencing: A review of automation strategies
J. F. Hess, T. A. Kohl, M. Kotrová, K. Rönsch, T. Paprotka, V. Mohr, T. Hutzenlaub, M. Brüggemann, R. Zengerle, S. Niemann, N. Paust Biotechnology Advances (2020-07) https://doi.org/ggth2v
DOI: 10.1016/j.biotechadv.2020.107537 · PMID: 32199980
341. Diagnosing COVID-19: The Disease and Tools for Detection
Buddhisha Udugama, Pranav Kadhiresan, Hannah N. Kozlowski, Ayden Malekjahani, Matthew Osborne, Vanessa Y. C. Li, Hongmin Chen, Samira Mubareka, Jonathan B. Gubbay, Warren C. W. Chan ACS Nano (2020-03-30) https://doi.org/ggq8ds
DOI: 10.1021/acsnano.0c02624 · PMID: 32223179 · PMCID: PMC7144809
342. Molecular Diagnosis of a Novel Coronavirus (2019-nCoV) Causing an Outbreak of Pneumonia
Daniel KW Chu, Yang Pan, Samuel MS Cheng, Kenrie PY Hui, Pavithra Krishnan, Yingzhi Liu, Daisy YM Ng, Carrie KC Wan, Peng Yang, Quanyi Wang, … Leo LM Poon Clinical Chemistry (2020-04) https://doi.org/ggnbpp
DOI: 10.1093/clinchem/hvaa029 · PMID: 32031583 · PMCID: PMC7108203
344. ddPCR: a more accurate tool for SARS-CoV-2 detection in low viral load specimens
Tao Suo, Xinjin Liu, Jiangpeng Feng, Ming Guo, Wenjia Hu, Dong Guo, Hafiz Ullah, Yang Yang, Qiuhan Zhang, Xin Wang, … Yu Chen Emerging Microbes & Infections (2020-06-07) https://doi.org/ggx2t2
DOI: 10.1080/22221751.2020.1772678 · PMID: 32438868 · PMCID: PMC7448897
345. Highly accurate and sensitive diagnostic detection of SARS-CoV-2 by digital PCR
Lianhua Dong, Junbo Zhou, Chunyan Niu, Quanyi Wang, Yang Pan, Sitong Sheng, Xia Wang, Yongzhuo Zhang, Jiayi Yang, Manqing Liu, … Xiang Fang Talanta (2021-03) https://doi.org/gh2jvj
DOI: 10.1016/j.talanta.2020.121726 · PMID: 33379001 · PMCID: PMC7588801
346. Evaluation of COVID-19 RT-qPCR test in multi-sample pools
Idan Yelin, Noga Aharony, Einat Shaer Tamar, Amir Argoetti, Esther Messer, Dina Berenbaum, Einat Shafran, Areen Kuzli, Nagam Gandali, Tamar Hashimshony, … Roy Kishony Cold Spring Harbor Laboratory (2020-03-27) https://doi.org/ggrn74
DOI: 10.1101/2020.03.26.20039438
347. Analytical Validation of a COVID-19 qRT-PCR Detection Assay Using a 384-well Format and Three Extraction Methods
Andrew C. Nelson, Benjamin Auch, Matthew Schomaker, Daryl M. Gohl, Patrick Grady, Darrell Johnson, Robyn Kincaid, Kylene E. Karnuth, Jerry Daniel, Jessica K. Fiege, … Sophia Yohe Cold Spring Harbor Laboratory (2020-04-05) https://doi.org/ggs45d
DOI: 10.1101/2020.04.02.022186
348. Nucleic acid detection with CRISPR-Cas13a/C2c2
Jonathan S. Gootenberg, Omar O. Abudayyeh, Jeong Wook Lee, Patrick Essletzbichler, Aaron J. Dy, Julia Joung, Vanessa Verdine, Nina Donghia, Nichole M. Daringer, Catherine A. Freije, … Feng Zhang Science (2017-04-28) https://doi.org/f93x8p
DOI: 10.1126/science.aam9321 · PMID: 28408723 · PMCID: PMC5526198
349. Development and Evaluation of A CRISPR-based Diagnostic For 2019-novel Coronavirus
Tieying Hou, Weiqi Zeng, Minling Yang, Wenjing Chen, Lili Ren, Jingwen Ai, Ji Wu, Yalong Liao, Xuejing Gou, Yongjun Li, … Teng Xu Cold Spring Harbor Laboratory (2020-02-25) https://doi.org/gg7km8
DOI: 10.1101/2020.02.22.20025460
350. CRISPR-based surveillance for COVID-19 using genomically-comprehensive machine learning design
Hayden C. Metsky, Catherine A. Freije, Tinna-Solveig F. Kosoko-Thoroddsen, Pardis C. Sabeti, Cameron Myhrvold Cold Spring Harbor Laboratory (2020-03-02) https://doi.org/ggr3zf
DOI: 10.1101/2020.02.26.967026
351. A Scalable, Easy-to-Deploy, Protocol for Cas13-Based Detection of SARS-CoV-2 Genetic Material
Jennifer N. Rauch, Eric Valois, Sabrina C. Solley, Friederike Braig, Ryan S. Lach, Morgane Audouard, Jose Carlos Ponce-Rojas, Michael S. Costello, Naomi J. Baxter, Kenneth S. Kosik, … Maxwell Z. Wilson Cold Spring Harbor Laboratory (2020-08-29) https://doi.org/gg7km7
DOI: 10.1101/2020.04.20.052159
352. CRISPR–Cas12-based detection of SARS-CoV-2
James P. Broughton, Xianding Deng, Guixia Yu, Clare L. Fasching, Venice Servellita, Jasmeet Singh, Xin Miao, Jessica A. Streithorst, Andrea Granados, Alicia Sotomayor-Gonzalez, … Charles Y. Chiu Nature Biotechnology (2020-04-16) https://doi.org/ggv47f
DOI: 10.1038/s41587-020-0513-4 · PMID: 32300245
353. An ultrasensitive, rapid, and portable coronavirus SARS-CoV-2 sequence detection method based on CRISPR-Cas12
Curti Lucia, Pereyra-Bonnet Federico, Gimenez Carla Alejandra Cold Spring Harbor Laboratory (2020-03-02) https://doi.org/gg7km6
DOI: 10.1101/2020.02.29.971127
354. All-in-One Dual CRISPR-Cas12a (AIOD-CRISPR) Assay: A Case for Rapid, Ultrasensitive and Visual Detection of Novel Coronavirus SARS-CoV-2 and HIV virus
Xiong Ding, Kun Yin, Ziyue Li, Changchun Liu Cold Spring Harbor Laboratory (2020-03-21) https://doi.org/gg7km5
DOI: 10.1101/2020.03.19.998724 · PMID: 32511323 · PMCID: PMC7239053
355. SARS-CoV-2 detection with CRISPR diagnostics
Lu Guo, Xuehan Sun, Xinge Wang, Chen Liang, Haiping Jiang, Qingqin Gao, Moyu Dai, Bin Qu, Sen Fang, Yihuan Mao, … Wei Li Cold Spring Harbor Laboratory (2020-04-11) https://doi.org/gg7kns
DOI: 10.1101/2020.04.10.023358
356. Electric-field-driven microfluidics for rapid CRISPR-based diagnostics and its application to detection of SARS-CoV-2
Ashwin Ramachandran, Diego A. Huyke, Eesha Sharma, Malaya K. Sahoo, Niaz Banaei, Benjamin A. Pinsky, Juan G. Santiago Cold Spring Harbor Laboratory (2020-05-22) https://doi.org/gg7knt
DOI: 10.1101/2020.05.21.109637
357. Rapid, sensitive and specific SARS coronavirus-2 detection: a multi-center comparison between standard qRT-PCR and CRISPR based DETECTR
Eelke Brandsma, Han J. M. P. Verhagen, Thijs J. W. van de Laar, Eric C. J. Claas, Marion Cornelissen, Emile van den Akker Cold Spring Harbor Laboratory (2020-07-29) https://doi.org/gg7km4
DOI: 10.1101/2020.07.27.20147249
359. The standard coronavirus test, if available, works well—but can new diagnostics help in this pandemic?
Robert Service Science (2020-03-22) https://doi.org/ggq9wm
DOI: 10.1126/science.abb8400
360. Laboratory Diagnosis of COVID-19: Current Issues and Challenges
Yi-Wei Tang, Jonathan E. Schmitz, David H. Persing, Charles W. Stratton Journal of Clinical Microbiology (2020-05-26) https://doi.org/ggq7h8
DOI: 10.1128/jcm.00512-20 · PMID: 32245835
361. Negative Nasopharyngeal and Oropharyngeal Swabs Do Not Rule Out COVID-19
Poramed Winichakoon, Romanee Chaiwarith, Chalerm Liwsrisakun, Parichat Salee, Aree Goonna, Atikun Limsukon, Quanhathai Kaewpoowat Journal of Clinical Microbiology (2020-04-23) https://doi.org/ggpw9m
DOI: 10.1128/jcm.00297-20 · PMID: 32102856 · PMCID: PMC7180262
362. A systematic review of antibody mediated immunity to coronaviruses: antibody kinetics, correlates of protection, and association of antibody responses with severity of disease
Angkana T. Huang, Bernardo Garcia-Carreras, Matt D. T. Hitchings, Bingyi Yang, Leah Katzelnick, Susan M Rattigan, Brooke Borgert, Carlos Moreno, Benjamin D. Solomon, Isabel Rodriguez-Barraquer, … Derek A. T. Cummings Cold Spring Harbor Laboratory (2020-04-17) https://doi.org/ggsfmz
DOI: 10.1101/2020.04.14.20065771 · PMID: 32511434
363. Longitudinal profile of antibodies against SARS-coronavirus in SARS patients and their clinical significance
Hongying MO, Guangqiao ZENG, Xiaolan REN, Hui LI, Changwen KE, Yaxia TAN, Chaoda CAI, Kefang LAI, Rongchang CHEN, Moira CHAN-YEUNG, Nanshan ZHONG Respirology (2006-01) https://doi.org/dn23vj
DOI: 10.1111/j.1440-1843.2006.00783.x · PMID: 16423201 · PMCID: PMC7192223
364. Two‐Year Prospective Study of the Humoral Immune Response of Patients with Severe Acute Respiratory Syndrome
Wei Liu, Arnaud Fontanet, Pan‐He Zhang, Lin Zhan, Zhong‐Tao Xin, Laurence Baril, Fang Tang, Hui Lv, Wu‐Chun Cao The Journal of Infectious Diseases (2006-03-15) https://doi.org/cmzn2k
DOI: 10.1086/500469 · PMID: 16479513 · PMCID: PMC7109932
365. The time course of the immune response to experimental coronavirus infection of man
K. A. Callow, H. F. Parry, M. Sergeant, D. A. J. Tyrrell Epidemiology and Infection (2009-05-15) https://doi.org/c9pnmg
DOI: 10.1017/s0950268800048019 · PMID: 2170159 · PMCID: PMC2271881
366. Loss of Bcl-6-Expressing T Follicular Helper Cells and Germinal Centers in COVID-19
Naoki Kaneko, Hsiao-Hsuan Kuo, Julie Boucau, Jocelyn R. Farmer, Hugues Allard-Chamard, Vinay S. Mahajan, Alicja Piechocka-Trocha, Kristina Lefteri, Matthew Osborn, Julia Bals, … Shiv Pillai Cell (2020-10) https://doi.org/gg9rdv
DOI: 10.1016/j.cell.2020.08.025 · PMID: 32877699 · PMCID: PMC7437499
367. A Peptide-Based Magnetic Chemiluminescence Enzyme Immunoassay for Serological Diagnosis of Coronavirus Disease 2019
Xue-fei Cai, Juan Chen, Jie- li Hu, Quan-xin Long, Hai-jun Deng, Ping Liu, Kai Fan, Pu Liao, Bei-zhong Liu, Gui-cheng Wu, … De-qiang Wang The Journal of Infectious Diseases (2020-07-15) https://doi.org/ggv2fx
DOI: 10.1093/infdis/jiaa243 · PMID: 32382737 · PMCID: PMC7239108
368. Deployment of convalescent plasma for the prevention and treatment of COVID-19
Evan M. Bloch, Shmuel Shoham, Arturo Casadevall, Bruce S. Sachais, Beth Shaz, Jeffrey L. Winters, Camille van Buskirk, Brenda J. Grossman, Michael Joyner, Jeffrey P. Henderson, … Aaron A. R. Tobian Journal of Clinical Investigation (2020-06-01) https://doi.org/ggr2w6
DOI: 10.1172/jci138745 · PMID: 32254064 · PMCID: PMC7259988
369. Treatment of 5 Critically Ill Patients With COVID-19 With Convalescent Plasma
Chenguang Shen, Zhaoqin Wang, Fang Zhao, Yang Yang, Jinxiu Li, Jing Yuan, Fuxiang Wang, Delin Li, Minghui Yang, Li Xing, … Lei Liu JAMA (2020-04-28) https://doi.org/dqn7
DOI: 10.1001/jama.2020.4783 · PMID: 32219428 · PMCID: PMC7101507
370. Convalescent Plasma Antibody Levels and the Risk of Death from Covid-19
Michael J. Joyner, Rickey E. Carter, Jonathon W. Senefeld, Stephen A. Klassen, John R. Mills, Patrick W. Johnson, Elitza S. Theel, Chad C. Wiggins, Katelyn A. Bruno, Allan M. Klompas, … Arturo Casadevall New England Journal of Medicine (2021-01-13) https://doi.org/ghs26g
DOI: 10.1056/nejmoa2031893 · PMID: 33523609 · PMCID: PMC7821984
371. Antibody Responses 8 Months after Asymptomatic or Mild SARS-CoV-2 Infection
Pyoeng Gyun Choe, Kye-Hyung Kim, Chang Kyung Kang, Hyeon Jeong Suh, EunKyo Kang, Sun Young Lee, Nam Joong Kim, Jongyoun Yi, Wan Beom Park, Myoung-don Oh Emerging Infectious Diseases (2021-03) https://doi.org/ghs9kq
DOI: 10.3201/eid2703.204543 · PMID: 33350923
372. Immunological memory to SARS-CoV-2 assessed for up to 8 months after infection
Jennifer M. Dan, Jose Mateus, Yu Kato, Kathryn M. Hastie, Esther Dawen Yu, Caterina E. Faliti, Alba Grifoni, Sydney I. Ramirez, Sonya Haupt, April Frazier, … Shane Crotty Science (2021-01-06) https://doi.org/ghrv9b
DOI: 10.1126/science.abf4063 · PMID: 33408181
373. Rapid generation of durable B cell memory to SARS-CoV-2 spike and nucleocapsid proteins in COVID-19 and convalescence.
Gemma E Hartley, Emily SJ Edwards, Pei M Aui, Nirupama Varese, Stephanie Stojanovic, James McMahon, Anton Y Peleg, Irene Boo, Heidi E Drummer, P Mark Hogarth, … Menno C van Zelm Science immunology (2020-12-22) https://www.ncbi.nlm.nih.gov/pubmed/33443036
DOI: 10.1126/sciimmunol.abf8891 · PMID: 33443036
374. Persistence of SARS-CoV-2-specific B and T cell responses in convalescent COVID-19 patients 6–8 months after the infection
Natalia Sherina, Antonio Piralla, Likun Du, Hui Wan, Makiko Kumagai-Braesch, Juni Andréll, Sten Braesch-Andersen, Irene Cassaniti, Elena Percivalle, Antonella Sarasini, … Qiang Pan-Hammarström Med (2021-03) https://doi.org/gh3xkz
DOI: 10.1016/j.medj.2021.02.001 · PMID: 33589885 · PMCID: PMC7874960
375. Evolution of antibody immunity to SARS-CoV-2
Christian Gaebler, Zijun Wang, Julio C. C. Lorenzi, Frauke Muecksch, Shlomo Finkin, Minami Tokuyama, Alice Cho, Mila Jankovic, Dennis Schaefer-Babajew, Thiago Y. Oliveira, … Michel C. Nussenzweig Nature (2021-01-18) https://doi.org/fq6k
DOI: 10.1038/s41586-021-03207-w · PMID: 33461210
376. Robust neutralizing antibodies to SARS-CoV-2 infection persist for months
Ania Wajnberg, Fatima Amanat, Adolfo Firpo, Deena R. Altman, Mark J. Bailey, Mayce Mansour, Meagan McMahon, Philip Meade, Damodara Rao Mendu, Kimberly Muellers, … Carlos Cordon-Cardo Science (2020-12-04) https://doi.org/fgfs
DOI: 10.1126/science.abd7728 · PMID: 33115920 · PMCID: PMC7810037
377. Longitudinal observation and decline of neutralizing antibody responses in the three months following SARS-CoV-2 infection in humans
Jeffrey Seow, Carl Graham, Blair Merrick, Sam Acors, Suzanne Pickering, Kathryn J. A. Steel, Oliver Hemmings, Aoife O’Byrne, Neophytos Kouphou, Rui Pedro Galao, … Katie J. Doores Nature Microbiology (2020-10-26) https://doi.org/fh7j
DOI: 10.1038/s41564-020-00813-8 · PMID: 33106674
378. Evolution of immune responses to SARS-CoV-2 in mild-moderate COVID-19
Adam K. Wheatley, Jennifer A. Juno, Jing J. Wang, Kevin J. Selva, Arnold Reynaldi, Hyon-Xhi Tan, Wen Shi Lee, Kathleen M. Wragg, Hannah G. Kelly, Robyn Esterbauer, … Stephen J. Kent Nature Communications (2021-02-19) https://doi.org/gh9vd5
DOI: 10.1038/s41467-021-21444-5 · PMID: 33608522 · PMCID: PMC7896046
379. COVID-19-neutralizing antibodies predict disease severity and survival
Wilfredo F. Garcia-Beltran, Evan C. Lam, Michael G. Astudillo, Diane Yang, Tyler E. Miller, Jared Feldman, Blake M. Hauser, Timothy M. Caradonna, Kiera L. Clayton, Adam D. Nitido, … Alejandro B. Balazs Cell (2021-01) https://doi.org/gh9vdx
DOI: 10.1016/j.cell.2020.12.015 · PMID: 33412089 · PMCID: PMC7837114
380. Functional SARS-CoV-2-Specific Immune Memory Persists after Mild COVID-19
Lauren B. Rodda, Jason Netland, Laila Shehata, Kurt B. Pruner, Peter A. Morawski, Christopher D. Thouvenel, Kennidy K. Takehara, Julie Eggenberger, Emily A. Hemann, Hayley R. Waterman, … Marion Pepper Cell (2021-01) https://doi.org/ghs9kf
DOI: 10.1016/j.cell.2020.11.029 · PMID: 33296701 · PMCID: PMC7682481
381. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals
Alba Grifoni, Daniela Weiskopf, Sydney I. Ramirez, Jose Mateus, Jennifer M. Dan, Carolyn Rydyznski Moderbacher, Stephen A. Rawlings, Aaron Sutherland, Lakshmanane Premkumar, Ramesh S. Jadi, … Alessandro Sette Cell (2020-06) https://doi.org/ggzxz2
DOI: 10.1016/j.cell.2020.05.015 · PMID: 32473127 · PMCID: PMC7237901
382. Robust SARS-CoV-2-specific T-cell immunity is maintained at 6 months following primary infection
J Zuo, A Dowell, H Pearce, K Verma, HM Long, J Begum, F Aiano, Z Amin-Chowdhury, B Hallis, L Stapley, … P Moss Cold Spring Harbor Laboratory (2020-11-02) https://doi.org/ghhrps
DOI: 10.1101/2020.11.01.362319
383. Persistent Cellular Immunity to SARS-CoV-2 Infection
Gaëlle Breton, Pilar Mendoza, Thomas Hagglof, Thiago Y. Oliveira, Dennis Schaefer-Babajew, Christian Gaebler, Martina Turroja, Arlene Hurley, Marina Caskey, Michel C. Nussenzweig Cold Spring Harbor Laboratory (2020-12-09) https://doi.org/ghs9kk
DOI: 10.1101/2020.12.08.416636 · PMID: 33330867 · PMCID: PMC7743071
384. Genomic evidence for reinfection with SARS-CoV-2: a case study
Richard L Tillett, Joel R Sevinsky, Paul D Hartley, Heather Kerwin, Natalie Crawford, Andrew Gorzalski, Chris Laverdure, Subhash C Verma, Cyprian C Rossetto, David Jackson, … Mark Pandori The Lancet Infectious Diseases (2021-01) https://doi.org/ghfgkt
DOI: 10.1016/s1473-3099(20)30764-7 · PMID: 33058797 · PMCID: PMC7550103
385. Asymptomatic Reinfection in 2 Healthcare Workers From India With Genetically Distinct Severe Acute Respiratory Syndrome Coronavirus 2
Vivek Gupta, Rahul C Bhoyar, Abhinav Jain, Saurabh Srivastava, Rashmi Upadhayay, Mohamed Imran, Bani Jolly, Mohit Kumar Divakar, Disha Sharma, Paras Sehgal, … Sridhar Sivasubbu Clinical Infectious Diseases (2020-09-23) https://doi.org/d97d
DOI: 10.1093/cid/ciaa1451 · PMID: 32964927 · PMCID: PMC7543380
386. Coronavirus Disease 2019 (COVID-19) Re-infection by a Phylogenetically Distinct Severe Acute Respiratory Syndrome Coronavirus 2 Strain Confirmed by Whole Genome Sequencing
Kelvin Kai-Wang To, Ivan Fan-Ngai Hung, Jonathan Daniel Ip, Allen Wing-Ho Chu, Wan-Mui Chan, Anthony Raymond Tam, Carol Ho-Yan Fong, Shuofeng Yuan, Hoi-Wah Tsoi, Anthony Chin-Ki Ng, … Kwok-Yung Yuen Clinical Infectious Diseases (2020-08-25) https://doi.org/d7ds
DOI: 10.1093/cid/ciaa1275 · PMID: 32840608 · PMCID: PMC7499500
389. Will SARS-CoV-2 Infection Elicit Long-Lasting Protective or Sterilising Immunity? Implications for Vaccine Strategies (2020)
David S. Kim, Sarah Rowland-Jones, Ester Gea-Mallorquí Frontiers in Immunology (2020-12-09) https://doi.org/ghs9ks
DOI: 10.3389/fimmu.2020.571481 · PMID: 33362759 · PMCID: PMC7756008
391. Detection of antibodies against SARS‐CoV‐2 in patients with COVID‐19
Zhe Du, Fengxue Zhu, Fuzheng Guo, Bo Yang, Tianbing Wang Journal of Medical Virology (2020-04-10) https://doi.org/ggq7m2
DOI: 10.1002/jmv.25820 · PMID: 32243608
392. A serological assay to detect SARS-CoV-2 seroconversion in humans
Fatima Amanat, Daniel Stadlbauer, Shirin Strohmeier, Thi H. O. Nguyen, Veronika Chromikova, Meagan McMahon, Kaijun Jiang, Guha Asthagiri Arunkumar, Denise Jurczyszak, Jose Polanco, … Florian Krammer Cold Spring Harbor Laboratory (2020-04-16) https://doi.org/ggpn83
DOI: 10.1101/2020.03.17.20037713 · PMID: 32511441
394. Strong associations and moderate predictive value of early symptoms for SARS-CoV-2 test positivity among healthcare workers, the Netherlands, March 2020
Alma Tostmann, John Bradley, Teun Bousema, Wing-Kee Yiek, Minke Holwerda, Chantal Bleeker-Rovers, Jaap ten Oever, Corianne Meijer, Janette Rahamat-Langendoen, Joost Hopman, … Heiman Wertheim Eurosurveillance (2020-04-23) https://doi.org/ggthwx
DOI: 10.2807/1560-7917.es.2020.25.16.2000508 · PMID: 32347200 · PMCID: PMC7189649
395. Correlation of Chest CT and RT-PCR Testing for Coronavirus Disease 2019 (COVID-19) in China: A Report of 1014 Cases
Tao Ai, Zhenlu Yang, Hongyan Hou, Chenao Zhan, Chong Chen, Wenzhi Lv, Qian Tao, Ziyong Sun, Liming Xia Radiology (2020-08) https://doi.org/ggmw6p
DOI: 10.1148/radiol.2020200642 · PMID: 32101510
396. Performance of Radiologists in Differentiating COVID-19 from Non-COVID-19 Viral Pneumonia at Chest CT
Harrison X. Bai, Ben Hsieh, Zeng Xiong, Kasey Halsey, Ji Whae Choi, Thi My Linh Tran, Ian Pan, Lin-Bo Shi, Dong-Cui Wang, Ji Mei, … Wei-Hua Liao Radiology (2020-08) https://doi.org/ggnqw4
DOI: 10.1148/radiol.2020200823 · PMID: 32155105
397. Covid-19: automatic detection from X-ray images utilizing transfer learning with convolutional neural networks
Ioannis D. Apostolopoulos, Tzani A. Mpesiana Physical and Engineering Sciences in Medicine (2020-04-03) https://doi.org/ggs448
DOI: 10.1007/s13246-020-00865-4 · PMCID: PMC7118364
403. Isolation of a Novel Coronavirus from a Man with Pneumonia in Saudi Arabia
Ali M. Zaki, Sander van Boheemen, Theo M. Bestebroer, Albert D. M. E. Osterhaus, Ron A. M. Fouchier New England Journal of Medicine (2012-11-08) https://doi.org/f4czx5
DOI: 10.1056/nejmoa1211721 · PMID: 23075143
404. Causes of Death and Comorbidities in Patients with COVID-19
Sefer Elezkurtaj, Selina Greuel, Jana Ihlow, Edward Michaelis, Philip Bischoff, Catarina Alisa Kunze, Bruno Valentin Sinn, Manuela Gerhold, Kathrin Hauptmann, Barbara Ingold-Heppner, … David Horst Cold Spring Harbor Laboratory (2020-06-17) https://doi.org/gg926j
DOI: 10.1101/2020.06.15.20131540
413. In Vitro and In Vivo Activities of Anti-Influenza Virus Compound T-705
Y. Furuta, K. Takahashi, Y. Fukuda, M. Kuno, T. Kamiyama, K. Kozaki, N. Nomura, H. Egawa, S. Minami, Y. Watanabe, … K. Shiraki Antimicrobial Agents and Chemotherapy (2002-04) https://doi.org/cndw7n
DOI: 10.1128/aac.46.4.977-981.2002 · PMID: 11897578 · PMCID: PMC127093
414. Efficacy of Orally Administered T-705 on Lethal Avian Influenza A (H5N1) Virus Infections in Mice
Robert W. Sidwell, Dale L. Barnard, Craig W. Day, Donald F. Smee, Kevin W. Bailey, Min-Hui Wong, John D. Morrey, Yousuke Furuta Antimicrobial Agents and Chemotherapy (2007-03) https://doi.org/dm9xr2
DOI: 10.1128/aac.01051-06 · PMID: 17194832 · PMCID: PMC1803113
415. Favipiravir (T-705), a broad spectrum inhibitor of viral RNA polymerase
Yousuke FURUTA, Takashi KOMENO, Takaaki NAKAMURA Proceedings of the Japan Academy, Series B (2017) https://doi.org/gbxcxw
DOI: 10.2183/pjab.93.027 · PMID: 28769016 · PMCID: PMC5713175
417. Activity of T-705 in a Hamster Model of Yellow Fever Virus Infection in Comparison with That of a Chemically Related Compound, T-1106
Justin G. Julander, Kristiina Shafer, Donald F. Smee, John D. Morrey, Yousuke Furuta Antimicrobial Agents and Chemotherapy (2009-01) https://doi.org/brknds
DOI: 10.1128/aac.01074-08 · PMID: 18955536 · PMCID: PMC2612161
418. In Vitro and In Vivo Activities of T-705 against Arenavirus and Bunyavirus Infections
Brian B. Gowen, Min-Hui Wong, Kie-Hoon Jung, Andrew B. Sanders, Michelle Mendenhall, Kevin W. Bailey, Yousuke Furuta, Robert W. Sidwell Antimicrobial Agents and Chemotherapy (2007-09) https://doi.org/d98c87
DOI: 10.1128/aac.00356-07 · PMID: 17606691 · PMCID: PMC2043187
419. Favipiravir (T-705) inhibits in vitro norovirus replication
J. Rocha-Pereira, D. Jochmans, K. Dallmeier, P. Leyssen, M. S. J. Nascimento, J. Neyts Biochemical and Biophysical Research Communications (2012-08) https://doi.org/f369j7
DOI: 10.1016/j.bbrc.2012.07.034 · PMID: 22809499
420. T-705 (Favipiravir) Inhibition of Arenavirus Replication in Cell Culture
Michelle Mendenhall, Andrew Russell, Terry Juelich, Emily L. Messina, Donald F. Smee, Alexander N. Freiberg, Michael R. Holbrook, Yousuke Furuta, Juan-Carlos de la Torre, Jack H. Nunberg, Brian B. Gowen Antimicrobial Agents and Chemotherapy (2011-02) https://doi.org/cppwsc
DOI: 10.1128/aac.01219-10 · PMID: 21115797 · PMCID: PMC3028760
421. The evolution of nucleoside analogue antivirals: A review for chemists and non-chemists. Part 1: Early structural modifications to the nucleoside scaffold
Katherine L. Seley-Radtke, Mary K. Yates Antiviral Research (2018-06) https://doi.org/gdpn35
DOI: 10.1016/j.antiviral.2018.04.004 · PMID: 29649496 · PMCID: PMC6396324
422. The Ambiguous Base-Pairing and High Substrate Efficiency of T-705 (Favipiravir) Ribofuranosyl 5′-Triphosphate towards Influenza A Virus Polymerase
Zhinan Jin, Lucas K. Smith, Vivek K. Rajwanshi, Baek Kim, Jerome Deval PLoS ONE (2013-07-10) https://doi.org/f5br92
DOI: 10.1371/journal.pone.0068347 · PMID: 23874596 · PMCID: PMC3707847
423. Experimental Treatment with Favipiravir for COVID-19: An Open-Label Control Study
Qingxian Cai, Minghui Yang, Dongjing Liu, Jun Chen, Dan Shu, Junxia Xia, Xuejiao Liao, Yuanbo Gu, Qiue Cai, Yang Yang, … Lei Liu Engineering (2020-10) https://doi.org/ggpprd
DOI: 10.1016/j.eng.2020.03.007 · PMID: 32346491
424. Repurposed Antiviral Drugs for Covid-19 — Interim WHO Solidarity Trial Results
WHO Solidarity Trial Consortium New England Journal of Medicine (2020-12-02) https://doi.org/ghnhnw
DOI: 10.1056/nejmoa2023184 · PMID: 33264556 · PMCID: PMC7727327
425. Lopinavir–ritonavir in patients admitted to hospital with COVID-19 (RECOVERY): a randomised, controlled, open-label, platform trial
Peter W Horby, Marion Mafham, Jennifer L Bell, Louise Linsell, Natalie Staplin, Jonathan Emberson, Adrian Palfreeman, Jason Raw, Einas Elmahi, Benjamin Prudon, … Martin J Landray The Lancet (2020-10) https://doi.org/fnx2
DOI: 10.1016/s0140-6736(20)32013-4 · PMID: 33031764 · PMCID: PMC7535623
426. A Large, Simple Trial Leading to Complex Questions
David P. Harrington, Lindsey R. Baden, Joseph W. Hogan New England Journal of Medicine (2020-12-02) https://doi.org/ghnhnx
DOI: 10.1056/nejme2034294 · PMID: 33264557 · PMCID: PMC7727323
428. Clinical Outcomes and Plasma Concentrations of Baloxavir Marboxil and Favipiravir in COVID-19 Patients: An Exploratory Randomized, Controlled Trial
Yan Lou, Lin Liu, Hangping Yao, Xingjiang Hu, Junwei Su, Kaijin Xu, Rui Luo, Xi Yang, Lingjuan He, Xiaoyang Lu, … Yunqing Qiu European Journal of Pharmaceutical Sciences (2021-02) https://doi.org/ghx88n
DOI: 10.1016/j.ejps.2020.105631 · PMID: 33115675 · PMCID: PMC7585719
429. Efficacy of favipiravir in COVID-19 treatment: a multi-center randomized study
Hany M. Dabbous, Sherief Abd-Elsalam, Manal H. El-Sayed, Ahmed F. Sherief, Fatma F. S. Ebeid, Mohamed Samir Abd El Ghafar, Shaimaa Soliman, Mohamed Elbahnasawy, Rehab Badawi, Mohamed Awad Tageldin Archives of Virology (2021-01-25) https://doi.org/ghx874
DOI: 10.1007/s00705-021-04956-9 · PMID: 33492523 · PMCID: PMC7829645
430. AVIFAVIR for Treatment of Patients With Moderate Coronavirus Disease 2019 (COVID-19): Interim Results of a Phase II/III Multicenter Randomized Clinical Trial
Andrey A Ivashchenko, Kirill A Dmitriev, Natalia V Vostokova, Valeria N Azarova, Andrew A Blinow, Alina N Egorova, Ivan G Gordeev, Alexey P Ilin, Ruben N Karapetian, Dmitry V Kravchenko, … Alexandre V Ivachtchenko Clinical Infectious Diseases (2020-08-09) https://doi.org/ghx9c2
DOI: 10.1093/cid/ciaa1176 · PMID: 32770240 · PMCID: PMC7454388
431. A review of the safety of favipiravir – a potential treatment in the COVID-19 pandemic?
Victoria Pilkington, Toby Pepperrell, Andrew Hill Journal of Virus Eradication (2020-04) https://doi.org/ftgm
DOI: 10.1016/s2055-6640(20)30016-9 · PMID: 32405421 · PMCID: PMC7331506
433. A Phase 3 Randomized Study to Evaluate the Safety and Antiviral Activity of Remdesivir (GS-5734™) in Participants With Severe COVID-19
Gilead Sciences clinicaltrials.gov (2020-12-15) https://clinicaltrials.gov/ct2/show/NCT04292899
434. A Multicenter, Adaptive, Randomized Blinded Controlled Trial of the Safety and Efficacy of Investigational Therapeutics for the Treatment of COVID-19 in Hospitalized Adults
National Institute of Allergy and Infectious Diseases (NIAID) clinicaltrials.gov (2020-12-05) https://clinicaltrials.gov/ct2/show/NCT04280705
435. Remdesivir for the Treatment of Covid-19 — Final Report
John H. Beigel, Kay M. Tomashek, Lori E. Dodd, Aneesh K. Mehta, Barry S. Zingman, Andre C. Kalil, Elizabeth Hohmann, Helen Y. Chu, Annie Luetkemeyer, Susan Kline, … H. Clifford Lane New England Journal of Medicine (2020-11-05) https://doi.org/dwkd
DOI: 10.1056/nejmoa2007764 · PMID: 32445440 · PMCID: PMC7262788
436. Compassionate Use of Remdesivir for Patients with Severe Covid-19
Jonathan Grein, Norio Ohmagari, Daniel Shin, George Diaz, Erika Asperges, Antonella Castagna, Torsten Feldt, Gary Green, Margaret L. Green, François-Xavier Lescure, … Timothy Flanigan New England Journal of Medicine (2020-06-11) https://doi.org/ggrm99
DOI: 10.1056/nejmoa2007016 · PMID: 32275812 · PMCID: PMC7169476
437. The antiviral compound remdesivir potently inhibits RNA-dependent RNA polymerase from Middle East respiratory syndrome coronavirus
Calvin J. Gordon, Egor P. Tchesnokov, Joy Y. Feng, Danielle P. Porter, Matthias Götte Journal of Biological Chemistry (2020-04) https://doi.org/ggqm6x
DOI: 10.1074/jbc.ac120.013056 · PMID: 32094225
438. Coronavirus Susceptibility to the Antiviral Remdesivir (GS-5734) Is Mediated by the Viral Polymerase and the Proofreading Exoribonuclease
Maria L. Agostini, Erica L. Andres, Amy C. Sims, Rachel L. Graham, Timothy P. Sheahan, Xiaotao Lu, Everett Clinton Smith, James Brett Case, Joy Y. Feng, Robert Jordan, … Mark R. Denison mBio (2018-03-06) https://doi.org/gc45v6
DOI: 10.1128/mbio.00221-18 · PMID: 29511076 · PMCID: PMC5844999
440. A Randomized, Controlled Trial of Ebola Virus Disease Therapeutics
Sabue Mulangu, Lori E. Dodd, Richard T. Davey, Olivier Tshiani Mbaya, Michael Proschan, Daniel Mukadi, Mariano Lusakibanza Manzo, Didier Nzolo, Antoine Tshomba Oloma, Augustin Ibanda, … the PALM Writing Group New England Journal of Medicine (2019-12-12) https://doi.org/ggqmx4
DOI: 10.1056/nejmoa1910993 · PMID: 31774950
441. Broad-spectrum antiviral GS-5734 inhibits both epidemic and zoonotic coronaviruses
Timothy P. Sheahan, Amy C. Sims, Rachel L. Graham, Vineet D. Menachery, Lisa E. Gralinski, James B. Case, Sarah R. Leist, Krzysztof Pyrc, Joy Y. Feng, Iva Trantcheva, … Ralph S. Baric Science Translational Medicine (2017-06-28) https://doi.org/gc3grb
DOI: 10.1126/scitranslmed.aal3653 · PMID: 28659436 · PMCID: PMC5567817
443. First 12 patients with coronavirus disease 2019 (COVID-19) in the United States
Stephanie A. Kujawski, Karen K Wong, Jennifer P. Collins, Lauren Epstein, Marie E. Killerby, Claire M. Midgley, Glen R. Abedi, N. Seema Ahmed, Olivia Almendares, Francisco N. Alvarez, … The COVID-19 Investigation Team Cold Spring Harbor Laboratory (2020-03-12) https://doi.org/ggqm6z
DOI: 10.1101/2020.03.09.20032896
444. First Case of 2019 Novel Coronavirus in the United States
Michelle L. Holshue, Chas DeBolt, Scott Lindquist, Kathy H. Lofy, John Wiesman, Hollianne Bruce, Christopher Spitters, Keith Ericson, Sara Wilkerson, Ahmet Tural, … Satish K. Pillai New England Journal of Medicine (2020-03-05) https://doi.org/ggjvr6
DOI: 10.1056/nejmoa2001191 · PMID: 32004427 · PMCID: PMC7092802
445. Remdesivir for 5 or 10 Days in Patients with Severe Covid-19
Jason D. Goldman, David C. B. Lye, David S. Hui, Kristen M. Marks, Raffaele Bruno, Rocio Montejano, Christoph D. Spinner, Massimo Galli, Mi-Young Ahn, Ronald G. Nahass, … Aruna Subramanian New England Journal of Medicine (2020-11-05) https://doi.org/ggz7qv
DOI: 10.1056/nejmoa2015301 · PMID: 32459919 · PMCID: PMC7377062
446. A Phase 3 Randomized Study to Evaluate the Safety and Antiviral Activity of Remdesivir (GS-5734™) in Participants With Moderate COVID-19 Compared to Standard of Care Treatment
Gilead Sciences clinicaltrials.gov (2021-01-21) https://clinicaltrials.gov/ct2/show/NCT04292730
449. A Phase 3 Randomized, Double-blind, Placebo-controlled, Multicenter Study to Evaluate the Efficacy and Safety of Remdesivir in Hospitalized Adult Patients With Severe COVID-19.
Bin Cao clinicaltrials.gov (2020-04-13) https://clinicaltrials.gov/ct2/show/NCT04257656
452. Conflicting results on the efficacy of remdesivir in hospitalized Covid-19 patients: comment on the Adaptive Covid-19 Treatment Trial
Leonarda Galiuto, Carlo Patrono European Heart Journal (2020-12-07) https://doi.org/ghp4kw
DOI: 10.1093/eurheartj/ehaa934 · PMID: 33306101 · PMCID: PMC7799042
454. Effect of Remdesivir vs Standard Care on Clinical Status at 11 Days in Patients With Moderate COVID-19
Christoph D. Spinner, Robert L. Gottlieb, Gerard J. Criner, José Ramón Arribas López, Anna Maria Cattelan, Alex Soriano Viladomiu, Onyema Ogbuagu, Prashant Malhotra, Kathleen M. Mullane, Antonella Castagna, … for the GS-US-540-5774 Investigators JAMA (2020-09-15) https://doi.org/ghhz6g
DOI: 10.1001/jama.2020.16349 · PMID: 32821939 · PMCID: PMC7442954
455. Baricitinib plus Remdesivir for Hospitalized Adults with Covid-19
Andre C. Kalil, Thomas F. Patterson, Aneesh K. Mehta, Kay M. Tomashek, Cameron R. Wolfe, Varduhi Ghazaryan, Vincent C. Marconi, Guillermo M. Ruiz-Palacios, Lanny Hsieh, Susan Kline, … John H. Beigel New England Journal of Medicine (2020-12-11) https://doi.org/ghpbd2
DOI: 10.1056/nejmoa2031994 · PMID: 33306283 · PMCID: PMC7745180
456. Letter of Authorization: EUA for baricitinib (Olumiant), in combination with remdesivir (Veklury), for the treatment of suspected or laboratory confirmed coronavirus disease 2019 (COVID-19)
Denise M. Hinton Food and Drug Administration (2020-01-19) https://www.fda.gov/media/143822/download
457. Proteases Essential for Human Influenza Virus Entry into Cells and Their Inhibitors as Potential Therapeutic Agents
Hiroshi Kido, Yuushi Okumura, Hiroshi Yamada, Trong Quang Le, Mihiro Yano Current Pharmaceutical Design (2007-02-01) https://doi.org/bts3xp
DOI: 10.2174/138161207780162971 · PMID: 17311557
458. Protease inhibitors targeting coronavirus and filovirus entry
Yanchen Zhou, Punitha Vedantham, Kai Lu, Juliet Agudelo, Ricardo Carrion, Jerritt W. Nunneley, Dale Barnard, Stefan Pöhlmann, James H. McKerrow, Adam R. Renslo, Graham Simmons Antiviral Research (2015-04) https://doi.org/ggr984
DOI: 10.1016/j.antiviral.2015.01.011 · PMID: 25666761 · PMCID: PMC4774534
459. Structure of Mpro from SARS-CoV-2 and discovery of its inhibitors
Zhenming Jin, Xiaoyu Du, Yechun Xu, Yongqiang Deng, Meiqin Liu, Yao Zhao, Bing Zhang, Xiaofeng Li, Leike Zhang, Chao Peng, … Haitao Yang Nature (2020-04-09) https://doi.org/ggrp42
DOI: 10.1038/s41586-020-2223-y · PMID: 32272481
461. The newly emerged SARS-Like coronavirus HCoV-EMC also has an “Achilles’ heel”: current effective inhibitor targeting a 3C-like protease
Zhilin Ren, Liming Yan, Ning Zhang, Yu Guo, Cheng Yang, Zhiyong Lou, Zihe Rao Protein & Cell (2013-04-03) https://doi.org/ggr7vh
DOI: 10.1007/s13238-013-2841-3 · PMID: 23549610 · PMCID: PMC4875521
462. Crystal structure of SARS-CoV-2 main protease provides a basis for design of improved α-ketoamide inhibitors
Linlin Zhang, Daizong Lin, Xinyuanyuan Sun, Ute Curth, Christian Drosten, Lucie Sauerhering, Stephan Becker, Katharina Rox, Rolf Hilgenfeld Science (2020-04-24) https://doi.org/ggp9sb
DOI: 10.1126/science.abb3405 · PMID: 32198291 · PMCID: PMC7164518
463. Ebselen, a promising antioxidant drug: mechanisms of action and targets of biological pathways
Gajendra Kumar Azad, Raghuvir S. Tomar Molecular Biology Reports (2014-05-28) https://doi.org/f6cnq3
DOI: 10.1007/s11033-014-3417-x · PMID: 24867080
464. Safety and efficacy of ebselen for the prevention of noise-induced hearing loss: a randomised, double-blind, placebo-controlled, phase 2 trial
Jonathan Kil, Edward Lobarinas, Christopher Spankovich, Scott K Griffiths, Patrick J Antonelli, Eric D Lynch, Colleen G Le Prell The Lancet (2017-09) https://doi.org/gbwnbv
DOI: 10.1016/s0140-6736(17)31791-9
466. Selenium Deficiency Is Associated with Mortality Risk from COVID-19
Arash Moghaddam, Raban Arved Heller, Qian Sun, Julian Seelig, Asan Cherkezov, Linda Seibert, Julian Hackler, Petra Seemann, Joachim Diegmann, Maximilian Pilz, … Lutz Schomburg Nutrients (2020-07-16) https://doi.org/gg5kbc
DOI: 10.3390/nu12072098 · PMID: 32708526 · PMCID: PMC7400921
467. Selenium and viral infection: are there lessons for COVID-19?
Giovanna Bermano, Catherine Méplan, Derry K. Mercer, John E. Hesketh British Journal of Nutrition (2020-08-06) https://doi.org/ghdx7w
DOI: 10.1017/s0007114520003128 · PMID: 32758306 · PMCID: PMC7503044
468. Selenium and selenoproteins in viral infection with potential relevance to COVID-19
Jinsong Zhang, Ramy Saad, Ethan Will Taylor, Margaret P. Rayman Redox Biology (2020-10) https://doi.org/ghdx7t
DOI: 10.1016/j.redox.2020.101715 · PMID: 32992282 · PMCID: PMC7481318
470. A Phase 2, Randomized, Double-Blind, Placebo-Controlled, Dose Escalation Study to Evaluate the Safety and Efficacy of SPI-1005 in Moderate COVID-19 Patients
Sound Pharmaceuticals, Incorporated clinicaltrials.gov (2021-03-24) https://clinicaltrials.gov/ct2/show/NCT04484025
471. A Phase 2, Randomized, Double-Blind, Placebo-Controlled, Dose Escalation Study to Evaluate the Safety and Efficacy of SPI-1005 in Severe COVID-19 Patients
Sound Pharmaceuticals, Incorporated clinicaltrials.gov (2021-03-24) https://clinicaltrials.gov/ct2/show/NCT04483973
472. Target discovery of ebselen with a biotinylated probe
Zhenzhen Chen, Zhongyao Jiang, Nan Chen, Qian Shi, Lili Tong, Fanpeng Kong, Xiufen Cheng, Hao Chen, Chu Wang, Bo Tang Chemical Communications (2018) https://doi.org/ggrtcm
DOI: 10.1039/c8cc04258f · PMID: 30091742
475. Hypertension Hot Potato — Anatomy of the Angiotensin-Receptor Blocker Recalls
J. Brian Byrd, Glenn M. Chertow, Vivek Bhalla New England Journal of Medicine (2019-04-25) https://doi.org/ggvc7g
DOI: 10.1056/nejmp1901657 · PMID: 30865819 · PMCID: PMC7066505
476. ACE Inhibitor and ARB Utilization and Expenditures in the Medicaid Fee-For-Service Program from 1991 to 2008
Boyang Bian, Christina M. L. Kelton, Jeff J. Guo, Patricia R. Wigle Journal of Managed Care Pharmacy (2010-11) https://doi.org/gh294c
DOI: 10.18553/jmcp.2010.16.9.671 · PMID: 21067253
477. Hypertension, the renin–angiotensin system, and the risk of lower respiratory tract infections and lung injury: implications for COVID-19
Reinhold Kreutz, Engi Abd El-Hady Algharably, Michel Azizi, Piotr Dobrowolski, Tomasz Guzik, Andrzej Januszewicz, Alexandre Persu, Aleksander Prejbisz, Thomas Günther Riemer, Ji-Guang Wang, Michel Burnier Cardiovascular Research (2020-08-01) https://doi.org/ggtwpj
DOI: 10.1093/cvr/cvaa097 · PMID: 32293003 · PMCID: PMC7184480
478. Angiotensin converting enzyme 2 activity and human atrial fibrillation: increased plasma angiotensin converting enzyme 2 activity is associated with atrial fibrillation and more advanced left atrial structural remodelling
Tomos E. Walters, Jonathan M. Kalman, Sheila K. Patel, Megan Mearns, Elena Velkoska, Louise M. Burrell Europace (2016-10-12) https://doi.org/gbt2jw
DOI: 10.1093/europace/euw246 · PMID: 27738071
479. Cardiovascular Disease, Drug Therapy, and Mortality in Covid-19
Mandeep R. Mehra, Sapan S. Desai, SreyRam Kuy, Timothy D. Henry, Amit N. Patel New England Journal of Medicine (2020-06-18) https://doi.org/ggtp6v
DOI: 10.1056/nejmoa2007621 · PMID: 32356626 · PMCID: PMC7206931
480. Retraction: Cardiovascular Disease, Drug Therapy, and Mortality in Covid-19. N Engl J Med. DOI: 10.1056/NEJMoa2007621.
Mandeep R. Mehra, Sapan S. Desai, SreyRam Kuy, Timothy D. Henry, Amit N. Patel New England Journal of Medicine (2020-06-25) https://doi.org/ggzkpj
DOI: 10.1056/nejmc2021225 · PMID: 32501665 · PMCID: PMC7274164
481. Response by Cohen et al to Letter Regarding Article, “Association of Inpatient Use of Angiotensin-Converting Enzyme Inhibitors and Angiotensin II Receptor Blockers With Mortality Among Patients With Hypertension Hospitalized With COVID-19”
Jordana B. Cohen, Thomas C. Hanff, Andrew M. South, Matthew A. Sparks, Swapnil Hiremath, Adam P. Bress, J. Brian Byrd, Julio A. Chirinos Circulation Research (2020-06-05) https://doi.org/gg3xsg
DOI: 10.1161/circresaha.120.317205 · PMID: 32496917 · PMCID: PMC7265880
482. Sound Science before Quick Judgement Regarding RAS Blockade in COVID-19
Matthew A. Sparks, Andrew South, Paul Welling, J. Matt Luther, Jordana Cohen, James Brian Byrd, Louise M. Burrell, Daniel Batlle, Laurie Tomlinson, Vivek Bhalla, … Swapnil Hiremath Clinical Journal of the American Society of Nephrology (2020-05-07) https://doi.org/ggq8gn
DOI: 10.2215/cjn.03530320 · PMID: 32220930 · PMCID: PMC7269218
489. Angiotensin Receptor Blockers and Angiotensin-converting Enzyme Inhibitors and Adverse Outcomes in Patients With COVID19 - Full Text View - ClinicalTrials.govhttps://clinicaltrials.gov/ct2/show/NCT04364893
490. Continuation versus discontinuation of renin–angiotensin system inhibitors in patients admitted to hospital with COVID-19: a prospective, randomised, open-label trial
Jordana B Cohen, Thomas C Hanff, Preethi William, Nancy Sweitzer, Nelson R Rosado-Santander, Carola Medina, Juan E Rodriguez-Mori, Nicolás Renna, Tara I Chang, Vicente Corrales-Medina, … Julio A Chirinos The Lancet Respiratory Medicine (2021-03) https://doi.org/fvgt
DOI: 10.1016/s2213-2600(20)30558-0 · PMID: 33422263 · PMCID: PMC7832152
491. Effect of Discontinuing vs Continuing Angiotensin-Converting Enzyme Inhibitors and Angiotensin II Receptor Blockers on Days Alive and Out of the Hospital in Patients Admitted With COVID-19
Renato D. Lopes, Ariane V. S. Macedo, Pedro G. M. de Barros E Silva, Renata J. Moll-Bernardes, Tiago M. dos Santos, Lilian Mazza, André Feldman, Guilherme D’Andréa Saba Arruda, Denílson C. de Albuquerque, Angelina S. Camiletti, … BRACE CORONA Investigators JAMA (2021-01-19) https://doi.org/gh2tw5
DOI: 10.1001/jama.2020.25864 · PMID: 33464336 · PMCID: PMC7816106
496. Chloroquine is a potent inhibitor of SARS coronavirus infection and spread
Martin J Vincent, Eric Bergeron, Suzanne Benjannet, Bobbie R Erickson, Pierre E Rollin, Thomas G Ksiazek, Nabil G Seidah, Stuart T Nichol Virology Journal (2005) https://doi.org/dvbds4
DOI: 10.1186/1743-422x-2-69 · PMID: 16115318 · PMCID: PMC1232869
497. In Vitro Antiviral Activity and Projection of Optimized Dosing Design of Hydroxychloroquine for the Treatment of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2)
Xueting Yao, Fei Ye, Miao Zhang, Cheng Cui, Baoying Huang, Peihua Niu, Xu Liu, Li Zhao, Erdan Dong, Chunli Song, … Dongyang Liu Clinical Infectious Diseases (2020-08-01) https://doi.org/ggpx7z
DOI: 10.1093/cid/ciaa237 · PMID: 32150618 · PMCID: PMC7108130
498. Mechanism of Action of Hydroxychloroquine in the Antiphospholipid Syndrome
Nadine Müller-Calleja, Davit Manukyan, Wolfram Ruf, Karl Lackner Blood (2016-12-02) https://doi.org/ggrm82
DOI: 10.1182/blood.v128.22.5023.5023
499. 14th International Congress on Antiphospholipid Antibodies Task Force Report on Antiphospholipid Syndrome Treatment Trends
Doruk Erkan, Cassyanne L. Aguiar, Danieli Andrade, Hannah Cohen, Maria J. Cuadrado, Adriana Danowski, Roger A. Levy, Thomas L. Ortel, Anisur Rahman, Jane E. Salmon, … Michael D. Lockshin Autoimmunity Reviews (2014-06) https://doi.org/ggp8r8
DOI: 10.1016/j.autrev.2014.01.053 · PMID: 24468415
501. COVID-19: a recommendation to examine the effect of hydroxychloroquine in preventing infection and progression
Dan Zhou, Sheng-Ming Dai, Qiang Tong Journal of Antimicrobial Chemotherapy (2020-07) https://doi.org/ggq84c
DOI: 10.1093/jac/dkaa114 · PMID: 32196083 · PMCID: PMC7184499
502. Hydroxychloroquine treatment of patients with human immunodeficiency virus type 1
Kirk Sperber, Michael Louie, Thomas Kraus, Jacqueline Proner, Erica Sapira, Su Lin, Vera Stecher, Lloyd Mayer Clinical Therapeutics (1995-07) https://doi.org/cq2hx9
DOI: 10.1016/0149-2918(95)80039-5
503. Hydroxychloroquine augments early virological response to pegylated interferon plus ribavirin in genotype-4 chronic hepatitis C patients
Gouda Kamel Helal, Magdy Abdelmawgoud Gad, Mohamed Fahmy Abd-Ellah, Mahmoud Saied Eid Journal of Medical Virology (2016-12) https://doi.org/f889nt
DOI: 10.1002/jmv.24575 · PMID: 27183377 · PMCID: PMC7167065
504. Hydroxychloroquine and azithromycin as a treatment of COVID-19: results of an open-label non-randomized clinical trial
Philippe Gautret, Jean-Christophe Lagier, Philippe Parola, Van Thuan Hoang, Line Meddeb, Morgane Mailhe, Barbara Doudier, Johan Courjon, Valérie Giordanengo, Vera Esteves Vieira, … Didier Raoult International Journal of Antimicrobial Agents (2020-07) https://doi.org/dp7d
DOI: 10.1016/j.ijantimicag.2020.105949 · PMID: 32205204 · PMCID: PMC7102549
507. No evidence of rapid antiviral clearance or clinical benefit with the combination of hydroxychloroquine and azithromycin in patients with severe COVID-19 infection
J. M. Molina, C. Delaugerre, J. Le Goff, B. Mela-Lima, D. Ponscarme, L. Goldwirt, N. de Castro Médecine et Maladies Infectieuses (2020-06) https://doi.org/ggqzrb
DOI: 10.1016/j.medmal.2020.03.006 · PMID: 32240719
508. Efficacy of hydroxychloroquine in patients with COVID-19: results of a randomized clinical trial
Zhaowei Chen, Jijia Hu, Zongwei Zhang, Shan Jiang, Shoumeng Han, Dandan Yan, Ruhong Zhuang, Ben Hu, Zhan Zhang Cold Spring Harbor Laboratory (2020-04-10) https://doi.org/ggqm4v
DOI: 10.1101/2020.03.22.20040758
511. A pilot study of hydroxychloroquine in treatment of patients with common coronavirus disease-19 (COVID-19)
CHEN Jun, LIU Danping, LIU Li, LIU Ping, XU Qingnian, XIA Lu, LING Yun, HUANG Dan, SONG Shuli, ZHANG Dandan, … LU Hongzhou Journal of Zhejiang University (Medical Sciences) (2020-03) https://doi.org/10.3785/j.issn.1008-9292.2020.03.03
DOI: 10.3785/j.issn.1008-9292.2020.03.03
512. Breakthrough: Chloroquine phosphate has shown apparent efficacy in treatment of COVID-19 associated pneumonia in clinical studies
Jianjun Gao, Zhenxue Tian, Xu Yang BioScience Trends (2020-02-29) https://doi.org/ggm3mv
DOI: 10.5582/bst.2020.01047 · PMID: 32074550
513. Targeting the Endocytic Pathway and Autophagy Process as a Novel Therapeutic Strategy in COVID-19
Naidi Yang, Han-Ming Shen International Journal of Biological Sciences (2020) https://doi.org/ggqspm
DOI: 10.7150/ijbs.45498 · PMID: 32226290 · PMCID: PMC7098027
515. RETRACTED: Hydroxychloroquine or chloroquine with or without a macrolide for treatment of COVID-19: a multinational registry analysis
Mandeep R Mehra, Sapan S Desai, Frank Ruschitzka, Amit N Patel The Lancet (2020-05) https://doi.org/ggwzsb
DOI: 10.1016/s0140-6736(20)31180-6 · PMID: 32450107 · PMCID: PMC7255293
516. Retraction—Hydroxychloroquine or chloroquine with or without a macrolide for treatment of COVID-19: a multinational registry analysis
Mandeep R Mehra, Frank Ruschitzka, Amit N Patel The Lancet (2020-06) https://doi.org/ggzqng
DOI: 10.1016/s0140-6736(20)31324-6 · PMID: 32511943 · PMCID: PMC7274621
517. Life Threatening Severe QTc Prolongation in Patient with Systemic Lupus Erythematosus due to Hydroxychloroquine
John P. O’Laughlin, Parag H. Mehta, Brian C. Wong Case Reports in Cardiology (2016) https://doi.org/ggqzrc
DOI: 10.1155/2016/4626279 · PMID: 27478650 · PMCID: PMC4960328
519. Safety of hydroxychloroquine, alone and in combination with azithromycin, in light of rapid wide-spread use for COVID-19: a multinational, network cohort and self-controlled case series study
Jennifer C.E.Lane, James Weaver, Kristin Kostka, Talita Duarte-Salles, Maria Tereza F. Abrahao, Heba Alghoul, Osaid Alser, Thamir M Alshammari, Patricia Biedermann, Edward Burn, … Daniel Prieto-Alhambra Cold Spring Harbor Laboratory (2020-04-10) https://doi.org/ggrn7s
DOI: 10.1101/2020.04.08.20054551
520. Chloroquine diphosphate in two different dosages as adjunctive therapy of hospitalized patients with severe respiratory syndrome in the context of coronavirus (SARS-CoV-2) infection: Preliminary safety results of a randomized, double-blinded, phase IIb clinical trial ( CloroCovid-19 Study )
Mayla Gabriela Silva Borba, Fernando Fonseca Almeida Val, Vanderson Souza Sampaio, Marcia Almeida Araújo Alexandre, Gisely Cardoso Melo, Marcelo Brito, Maria Paula Gomes Mourão, José Diego Brito-Sousa, Djane Baía-da-Silva, Marcus Vinitius Farias Guerra, … CloroCovid-19 Team Cold Spring Harbor Laboratory (2020-04-16) https://doi.org/ggr3nj
DOI: 10.1101/2020.04.07.20056424
523. Hydroxychloroquine in patients mainly with mild to moderate COVID–19: an open–label, randomized, controlled trial
Wei Tang, Zhujun Cao, Mingfeng Han, Zhengyan Wang, Junwen Chen, Wenjin Sun, Yaojie Wu, Wei Xiao, Shengyong Liu, Erzhen Chen, … Qing Xie Cold Spring Harbor Laboratory (2020-05-07) https://doi.org/ggr68m
DOI: 10.1101/2020.04.10.20060558
524. Outcomes of hydroxychloroquine usage in United States veterans hospitalized with Covid-19
Joseph Magagnoli, Siddharth Narendran, Felipe Pereira, Tammy Cummings, James W. Hardin, S. Scott Sutton, Jayakrishna Ambati Cold Spring Harbor Laboratory (2020-04-21) https://doi.org/ggspt6
DOI: 10.1101/2020.04.16.20065920 · PMID: 32511622 · PMCID: PMC7276049
526. Hydroxychloroquine for Early Treatment of Adults With Mild Coronavirus Disease 2019: A Randomized, Controlled Trial
Oriol Mitjà, Marc Corbacho-Monné, Maria Ubals, Cristian Tebé, Judith Peñafiel, Aurelio Tobias, Ester Ballana, Andrea Alemany, Núria Riera-Martí, Carla A Pérez, … Martí Vall-Mayans Clinical Infectious Diseases (2020-07-16) https://doi.org/gg5f9x
DOI: 10.1093/cid/ciaa1009 · PMID: 32674126 · PMCID: PMC7454406
527. A Randomized Trial of Hydroxychloroquine as Postexposure Prophylaxis for Covid-19
David R. Boulware, Matthew F. Pullen, Ananta S. Bangdiwala, Katelyn A. Pastick, Sarah M. Lofgren, Elizabeth C. Okafor, Caleb P. Skipper, Alanna A. Nascene, Melanie R. Nicol, Mahsa Abassi, … Kathy H. Hullsiek New England Journal of Medicine (2020-08-06) https://doi.org/dxkv
DOI: 10.1056/nejmoa2016638 · PMID: 32492293 · PMCID: PMC7289276
528. Efficacy and Safety of Hydroxychloroquine vs Placebo for Pre-exposure SARS-CoV-2 Prophylaxis Among Health Care Workers
Benjamin S. Abella, Eliana L. Jolkovsky, Barbara T. Biney, Julie E. Uspal, Matthew C. Hyman, Ian Frank, Scott E. Hensley, Saar Gill, Dan T. Vogl, Ivan Maillard, … Prevention and Treatment of COVID-19 With Hydroxychloroquine (PATCH) Investigators JAMA Internal Medicine (2021-02-01) https://doi.org/ghd6nj
DOI: 10.1001/jamainternmed.2020.6319 · PMID: 33001138 · PMCID: PMC7527945
529. Mechanisms of action of hydroxychloroquine and chloroquine: implications for rheumatology
Eva Schrezenmeier, Thomas Dörner Nature Reviews Rheumatology (2020-02-07) https://doi.org/ggzjnh
DOI: 10.1038/s41584-020-0372-x · PMID: 32034323
532. 16-METHYLATED STEROIDS. I. 16α-METHYLATED ANALOGS OF CORTISONE, A NEW GROUP OF ANTI-INFLAMMATORY STEROIDS
Glen E. Arth, David B. R. Johnston, John Fried, William W. Spooncer, Dale R. Hoff, Lewis H. Sarett Journal of the American Chemical Society (2002-05-01) https://doi.org/cj5c82
DOI: 10.1021/ja01545a061
535. Potential benefits of precise corticosteroids therapy for severe 2019-nCoV pneumonia
Wei Zhou, Yisi Liu, Dongdong Tian, Cheng Wang, Sa Wang, Jing Cheng, Ming Hu, Minghao Fang, Yue Gao Signal Transduction and Targeted Therapy (2020-02-21) https://doi.org/ggqr84
DOI: 10.1038/s41392-020-0127-9 · PMID: 32296012 · PMCID: PMC7035340
536. Dexamethasone treatment for the acute respiratory distress syndrome: a multicentre, randomised controlled trial
Jesús Villar, Carlos Ferrando, Domingo Martínez, Alfonso Ambrós, Tomás Muñoz, Juan A Soler, Gerardo Aguilar, Francisco Alba, Elena González-Higueras, Luís A Conesa, … Jesús Villar The Lancet Respiratory Medicine (2020-03) https://doi.org/ggpxzc
DOI: 10.1016/s2213-2600(19)30417-5
540. Non-traditional cytokines: How catecholamines and adipokines influence macrophages in immunity, metabolism and the central nervous system
Mark A. Barnes, Monica J. Carson, Meera G. Nair Cytokine (2015-04) https://doi.org/f65c59
DOI: 10.1016/j.cyto.2015.01.008 · PMID: 25703786 · PMCID: PMC4590987
542. Recovery of the Hypothalamic-Pituitary-Adrenal Response to Stress
Arantxa García, Octavi Martí, Astrid Vallès, Silvina Dal-Zotto, Antonio Armario Neuroendocrinology (2000) https://doi.org/b2cq8n
DOI: 10.1159/000054578 · PMID: 10971146
543. Modulatory effects of glucocorticoids and catecholamines on human interleukin-12 and interleukin-10 production: clinical implications.
IJ Elenkov, DA Papanicolaou, RL Wilder, GP Chrousos Proceedings of the Association of American Physicians (1996-09) https://www.ncbi.nlm.nih.gov/pubmed/8902882
PMID: 8902882
544. Prevention of infection caused by immunosuppressive drugs in gastroenterology
Katarzyna Orlicka, Eleanor Barnes, Emma L. Culver Therapeutic Advances in Chronic Disease (2013-04-22) https://doi.org/ggrqd3
DOI: 10.1177/2040622313485275 · PMID: 23819020 · PMCID: PMC3697844
546. Effect of Dexamethasone in Hospitalized Patients with COVID-19 – Preliminary Report
Peter Horby, Wei Shen Lim, Jonathan Emberson, Marion Mafham, Jennifer Bell, Louise Linsell, Natalie Staplin, Christopher Brightling, Andrew Ustianowski, Einas Elmahi, … RECOVERY Collaborative Group Cold Spring Harbor Laboratory (2020-06-22) https://doi.org/dz5x
DOI: 10.1101/2020.06.22.20137273
548. Corticosteroids for Patients With Coronavirus Disease 2019 (COVID-19) With Different Disease Severity: A Meta-Analysis of Randomized Clinical Trials
Laura Pasin, Paolo Navalesi, Alberto Zangrillo, Artem Kuzovlev, Valery Likhvantsev, Ludhmila Abrahão Hajjar, Stefano Fresilli, Marcus Vinicius Guimaraes Lacerda, Giovanni Landoni Journal of Cardiothoracic and Vascular Anesthesia (2021-02) https://doi.org/ghzkp9
DOI: 10.1053/j.jvca.2020.11.057 · PMID: 33298370 · PMCID: PMC7698829
549. Current concepts in the diagnosis and management of cytokine release syndrome
Daniel W. Lee, Rebecca Gardner, David L. Porter, Chrystal U. Louis, Nabil Ahmed, Michael Jensen, Stephan A. Grupp, Crystal L. Mackall Blood (2014-07-10) https://doi.org/ggsrwk
DOI: 10.1182/blood-2014-05-552729 · PMID: 24876563 · PMCID: PMC4093680
552. Dexamethasone for COVID-19: data needed from randomised clinical trials in Africa
Helen Brotherton, Effua Usuf, Behzad Nadjm, Karen Forrest, Kalifa Bojang, Ahmadou Lamin Samateh, Mustapha Bittaye, Charles AP Roberts, Umberto d’Alessandro, Anna Roca The Lancet Global Health (2020-09) https://doi.org/gg42kx
DOI: 10.1016/s2214-109x(20)30318-1 · PMID: 32679038 · PMCID: PMC7833918
555. Deriving Immune Modulating Drugs from Viruses—A New Class of Biologics
Jordan R. Yaron, Liqiang Zhang, Qiuyun Guo, Michelle Burgin, Lauren N. Schutz, Enkidia Awo, Lyn Wise, Kurt L. Krause, Cristhian J. Ildefonso, Jacek M. Kwiecien, … Alexandra R. Lucas Journal of Clinical Medicine (2020-03-31) https://doi.org/ghdx73
DOI: 10.3390/jcm9040972 · PMID: 32244484 · PMCID: PMC7230489
558. Hall of Fame among Pro-inflammatory Cytokines: Interleukin-6 Gene and Its Transcriptional Regulation Mechanisms
Yang Luo, Song Guo Zheng Frontiers in Immunology (2016-12-19) https://doi.org/ggqmgv
DOI: 10.3389/fimmu.2016.00604 · PMID: 28066415 · PMCID: PMC5165036
559. IL-6 Trans-Signaling via the Soluble IL-6 Receptor: Importance for the Pro-Inflammatory Activities of IL-6
Stefan Rose-John International Journal of Biological Sciences (2012) https://doi.org/f4c4hf
DOI: 10.7150/ijbs.4989 · PMID: 23136552 · PMCID: PMC3491447
561. Plasticity and cross-talk of Interleukin 6-type cytokines
Christoph Garbers, Heike M. Hermanns, Fred Schaper, Gerhard Müller-Newen, Joachim Grötzinger, Stefan Rose-John, Jürgen Scheller Cytokine & Growth Factor Reviews (2012-06) https://doi.org/f3z743
DOI: 10.1016/j.cytogfr.2012.04.001 · PMID: 22595692
562. Soluble receptors for cytokines and growth factors: generation and biological function
S Rose-John, PC Heinrich Biochemical Journal (1994-06-01) https://doi.org/ggqmgd
DOI: 10.1042/bj3000281 · PMID: 8002928 · PMCID: PMC1138158
563. Interleukin-6; pathogenesis and treatment of autoimmune inflammatory diseases
Toshio Tanaka, Masashi Narazaki, Kazuya Masuda, Tadamitsu Kishimoto Inflammation and Regeneration (2013) https://doi.org/ggqmgt
DOI: 10.2492/inflammregen.33.054
564. Effective treatment of severe COVID-19 patients with tocilizumab
Xiaoling Xu, Mingfeng Han, Tiantian Li, Wei Sun, Dongsheng Wang, Binqing Fu, Yonggang Zhou, Xiaohu Zheng, Yun Yang, Xiuyong Li, … Haiming Wei Proceedings of the National Academy of Sciences (2020-05-19) https://doi.org/ggv3r3
DOI: 10.1073/pnas.2005615117 · PMID: 32350134 · PMCID: PMC7245089
565. Tocilizumab in patients with severe COVID-19: a retrospective cohort study
Giovanni Guaraldi, Marianna Meschiari, Alessandro Cozzi-Lepri, Jovana Milic, Roberto Tonelli, Marianna Menozzi, Erica Franceschini, Gianluca Cuomo, Gabriella Orlando, Vanni Borghi, … Cristina Mussini The Lancet Rheumatology (2020-08) https://doi.org/d2pk
DOI: 10.1016/s2665-9913(20)30173-9 · PMID: 32835257 · PMCID: PMC7314456
566. Tocilizumab Treatment for Cytokine Release Syndrome in Hospitalized Patients With Coronavirus Disease 2019
Christina C. Price, Frederick L. Altice, Yu Shyr, Alan Koff, Lauren Pischel, George Goshua, Marwan M. Azar, Dayna Mcmanus, Sheau-Chiann Chen, Shana E. Gleeson, … Maricar Malinis Chest (2020-10) https://doi.org/gg2789
DOI: 10.1016/j.chest.2020.06.006 · PMID: 32553536 · PMCID: PMC7831876
567. Impact of low dose tocilizumab on mortality rate in patients with COVID-19 related pneumonia
Ruggero Capra, Nicola De Rossi, Flavia Mattioli, Giuseppe Romanelli, Cristina Scarpazza, Maria Pia Sormani, Stefania Cossi European Journal of Internal Medicine (2020-06) https://doi.org/ggx4fm
DOI: 10.1016/j.ejim.2020.05.009 · PMID: 32405160 · PMCID: PMC7219361
568. Tocilizumab therapy reduced intensive care unit admissions and/or mortality in COVID-19 patients
T. Klopfenstein, S. Zayet, A. Lohse, J.-C. Balblanc, J. Badie, P.-Y. Royer, L. Toko, C. Mezher, N. J. Kadiane-Oussou, M. Bossert, … T. Conrozier Médecine et Maladies Infectieuses (2020-08) https://doi.org/ggvz45
DOI: 10.1016/j.medmal.2020.05.001 · PMID: 32387320 · PMCID: PMC7202806
569. Outcomes in patients with severe COVID-19 disease treated with tocilizumab: a case–controlled study
G Rojas-Marte, M Khalid, O Mukhtar, AT Hashmi, MA Waheed, S Ehrlich, A Aslam, S Siddiqui, C Agarwal, Y Malyshev, … J Shani QJM: An International Journal of Medicine (2020-08) https://doi.org/gg496t
DOI: 10.1093/qjmed/hcaa206 · PMID: 32569363 · PMCID: PMC7337835
570. Systematic Review and Meta-Analysis of Case-Control Studies from 7,000 COVID-19 Pneumonia Patients Suggests a Beneficial Impact of Tocilizumab with Benefit Most Evident in Non-Corticosteroid Exposed Subjects.
Abdulla Watad, Nicola Luigi Bragazzi, Charlie Bridgewood, Muhammad Mansour, Naim Mahroum, Matteo Riccò, Ahmed Nasr, Amr Hussein, Omer Gendelman, Yehuda Shoenfeld, … Dennis McGonagle SSRN Electronic Journal (2020) https://doi.org/gg62hz
DOI: 10.2139/ssrn.3642653
571. The efficacy of IL-6 inhibitor Tocilizumab in reducing severe COVID-19 mortality: a systematic review
Avi Gurion Kaye, Robert Siegel PeerJ (2020-11-02) https://doi.org/ghx8r4
DOI: 10.7717/peerj.10322 · PMID: 33194450 · PMCID: PMC7643559
572. Rationale and evidence on the use of tocilizumab in COVID-19: a systematic review
A. Cortegiani, M. Ippolito, M. Greco, V. Granone, A. Protti, C. Gregoretti, A. Giarratano, S. Einav, M. Cecconi Pulmonology (2021-01) https://doi.org/gg5xv3
DOI: 10.1016/j.pulmoe.2020.07.003 · PMID: 32713784 · PMCID: PMC7369580
573. Tocilizumab in patients admitted to hospital with COVID-19 (RECOVERY): preliminary results of a randomised, controlled, open-label, platform trial
Peter W Horby, Guilherme Pessoa-Amorim, Leon Peto, Christopher E Brightling, Rahuldeb Sarkar, Koshy Thomas, Vandana Jeebun, Abdul Ashish, Redmond Tully, David Chadwick, … RECOVERY Collaborative Group Cold Spring Harbor Laboratory (2021-02-11) https://doi.org/fvqj
DOI: 10.1101/2021.02.11.21249258
575. Tocilizumab during pregnancy and lactation: drug levels in maternal serum, cord blood, breast milk and infant serum
Jumpei Saito, Naho Yakuwa, Kayoko Kaneko, Chinatsu Takai, Mikako Goto, Ken Nakajima, Akimasa Yamatani, Atsuko Murashima Rheumatology (2019-08) https://doi.org/ggzhks
DOI: 10.1093/rheumatology/kez100 · PMID: 30945743
576. Short-course tocilizumab increases risk of hepatitis B virus reactivation in patients with rheumatoid arthritis: a prospective clinical observation
Le-Feng Chen, Ying-Qian Mo, Jun Jing, Jian-Da Ma, Dong-Hui Zheng, Lie Dai International Journal of Rheumatic Diseases (2017-07) https://doi.org/f9pbc5
DOI: 10.1111/1756-185x.13010 · PMID: 28160426
578. Risk of adverse events including serious infections in rheumatoid arthritis patients treated with tocilizumab: a systematic literature review and meta-analysis of randomized controlled trials
L. Campbell, C. Chen, S. S. Bhagat, R. A. Parker, A. J. K. Ostor Rheumatology (2010-11-14) https://doi.org/crqn7c
DOI: 10.1093/rheumatology/keq343 · PMID: 21078627
579. Risk of serious infections in tocilizumab versus other biologic drugs in patients with rheumatoid arthritis: a multidatabase cohort study
Ajinkya Pawar, Rishi J Desai, Daniel H Solomon, Adrian J Santiago Ortiz, Sara Gale, Min Bao, Khaled Sarsour, Sebastian Schneeweiss, Seoyoung C Kim Annals of the Rheumatic Diseases (2019-04) https://doi.org/gg62hx
DOI: 10.1136/annrheumdis-2018-214367 · PMID: 30679153
580. Risk of infections in rheumatoid arthritis patients treated with tocilizumab
Veronika R. Lang, Matthias Englbrecht, Jürgen Rech, Hubert Nüsslein, Karin Manger, Florian Schuch, Hans-Peter Tony, Martin Fleck, Bernhard Manger, Georg Schett, Jochen Zwerina Rheumatology (2012-05) https://doi.org/d3b3rh
DOI: 10.1093/rheumatology/ker223 · PMID: 21865281
582. Incidence of ARDS and outcomes in hospitalized patients with COVID-19: a global literature survey
Susan J. Tzotzos, Bernhard Fischer, Hendrik Fischer, Markus Zeitlinger Critical Care (2020-08-21) https://doi.org/gh294r
DOI: 10.1186/s13054-020-03240-7 · PMID: 32825837 · PMCID: PMC7441837
583. The Efficacy of IL-6 Inhibitor Tocilizumab in Reducing Severe COVID-19 Mortality: A Systematic Review
Avi Kaye, Robert Siegel Cold Spring Harbor Laboratory (2020-07-14) https://doi.org/gg62hv
DOI: 10.1101/2020.07.10.20150938
584. Utilizing tocilizumab for the treatment of cytokine release syndrome in COVID-19
Ali Hassoun, Elizabeth Dilip Thottacherry, Justin Muklewicz, Qurrat-ul-ain Aziz, Jonathan Edwards Journal of Clinical Virology (2020-07) https://doi.org/ggx359
DOI: 10.1016/j.jcv.2020.104443 · PMID: 32425661 · PMCID: PMC7229471
585. Development of therapeutic antibodies for the treatment of diseases
Ruei-Min Lu, Yu-Chyi Hwang, I-Ju Liu, Chi-Chiu Lee, Han-Zen Tsai, Hsin-Jung Li, Han-Chung Wu Journal of Biomedical Science (2020-01-02) https://doi.org/ggqbpx
DOI: 10.1186/s12929-019-0592-z · PMID: 31894001 · PMCID: PMC6939334
591. Chronological evolution of IgM, IgA, IgG and neutralisation antibodies after infection with SARS-associated coronavirus
P.-R. Hsueh, L.-M. Huang, P.-J. Chen, C.-L. Kao, P.-C. Yang Clinical Microbiology and Infection (2004-12) https://doi.org/cwwg87
DOI: 10.1111/j.1469-0691.2004.01009.x · PMID: 15606632
592. Neutralizing Antibodies in Patients with Severe Acute Respiratory Syndrome-Associated Coronavirus Infection
Nie Yuchun, Wang Guangwen, Shi Xuanling, Zhang Hong, Qiu Yan, He Zhongping, Wang Wei, Lian Gewei, Yin Xiaolei, Du Liying, … Ding Mingxiao The Journal of Infectious Diseases (2004-09) https://doi.org/cgqj5b
DOI: 10.1086/423286 · PMID: 15319862
593. Potent human monoclonal antibodies against SARS CoV, Nipah and Hendra viruses
Ponraj Prabakaran, Zhongyu Zhu, Xiaodong Xiao, Arya Biragyn, Antony S Dimitrov, Christopher C Broder, Dimiter S Dimitrov Expert Opinion on Biological Therapy (2009-04-08) https://doi.org/b88kw8
DOI: 10.1517/14712590902763755 · PMID: 19216624 · PMCID: PMC2705284
594. SARS-CoV-2 and SARS-CoV Spike-RBD Structure and Receptor Binding Comparison and Potential Implications on Neutralizing Antibody and Vaccine Development
Chunyun Sun, Long Chen, Ji Yang, Chunxia Luo, Yanjing Zhang, Jing Li, Jiahui Yang, Jie Zhang, Liangzhi Xie Cold Spring Harbor Laboratory (2020-02-20) https://doi.org/ggq63j
DOI: 10.1101/2020.02.16.951723
595. Fruitful Neutralizing Antibody Pipeline Brings Hope To Defeat SARS-Cov-2
Alex Renn, Ying Fu, Xin Hu, Matthew D. Hall, Anton Simeonov Trends in Pharmacological Sciences (2020-11) https://doi.org/gg72sv
DOI: 10.1016/j.tips.2020.07.004 · PMID: 32829936 · PMCID: PMC7572790
596. Cross-neutralization of SARS-CoV-2 by a human monoclonal SARS-CoV antibody
Dora Pinto, Young-Jun Park, Martina Beltramello, Alexandra C. Walls, M. Alejandra Tortorici, Siro Bianchi, Stefano Jaconi, Katja Culap, Fabrizia Zatta, Anna De Marco, … Davide Corti Nature (2020-05-18) https://doi.org/dv4x
DOI: 10.1038/s41586-020-2349-y · PMID: 32422645
597. A human monoclonal antibody blocking SARS-CoV-2 infection
Chunyan Wang, Wentao Li, Dubravka Drabek, Nisreen M. A. Okba, Rien van Haperen, Albert D. M. E. Osterhaus, Frank J. M. van Kuppeveld, Bart L. Haagmans, Frank Grosveld, Berend-Jan Bosch Cold Spring Harbor Laboratory (2020-03-12) https://doi.org/ggnw4t
DOI: 10.1101/2020.03.11.987958
599. LY-CoV555, a rapidly isolated potent neutralizing antibody, provides protection in a non-human primate model of SARS-CoV-2 infection
Bryan E. Jones, Patricia L. Brown-Augsburger, Kizzmekia S. Corbett, Kathryn Westendorf, Julian Davies, Thomas P. Cujec, Christopher M. Wiethoff, Jamie L. Blackbourne, Beverly A. Heinz, Denisa Foster, … Ester Falconer Cold Spring Harbor Laboratory (2020-10-09) https://doi.org/gh4sjm
DOI: 10.1101/2020.09.30.318972 · PMID: 33024963 · PMCID: PMC7536866
601. A Phase 1, Randomized, Placebo-Controlled Study to Evaluate the Tolerability, Safety, Pharmacokinetics, and Immunogenicity of LY3832479 Given as a Single Intravenous Dose in Healthy Participants
Eli Lilly and Company clinicaltrials.gov (2020-10-07) https://clinicaltrials.gov/ct2/show/NCT04441931
602. Effect of Bamlanivimab as Monotherapy or in Combination With Etesevimab on Viral Load in Patients With Mild to Moderate COVID-19
Robert L. Gottlieb, Ajay Nirula, Peter Chen, Joseph Boscia, Barry Heller, Jason Morris, Gregory Huhn, Jose Cardona, Bharat Mocherla, Valentina Stosor, … Daniel M. Skovronsky JAMA (2021-02-16) https://doi.org/ghvnrr
DOI: 10.1001/jama.2021.0202 · PMID: 33475701 · PMCID: PMC7821080
603. A Study of LY3819253 (LY-CoV555) and LY3832479 (LY-CoV016) in Participants With Mild to Moderate COVID-19 Illness - Full Text View - ClinicalTrials.govhttps://clinicaltrials.gov/ct2/show/NCT04427501
604. SARS-CoV-2 Neutralizing Antibody LY-CoV555 in Outpatients with Covid-19
Peter Chen, Ajay Nirula, Barry Heller, Robert L. Gottlieb, Joseph Boscia, Jason Morris, Gregory Huhn, Jose Cardona, Bharat Mocherla, Valentina Stosor, … Daniel M. Skovronsky New England Journal of Medicine (2021-01-21) https://doi.org/fgtm
DOI: 10.1056/nejmoa2029849 · PMID: 33113295 · PMCID: PMC7646625
605. Studies in humanized mice and convalescent humans yield a SARS-CoV-2 antibody cocktail
Johanna Hansen, Alina Baum, Kristen E. Pascal, Vincenzo Russo, Stephanie Giordano, Elzbieta Wloga, Benjamin O. Fulton, Ying Yan, Katrina Koon, Krunal Patel, … Christos A. Kyratsous Science (2020-08-21) https://doi.org/fcqh
DOI: 10.1126/science.abd0827 · PMID: 32540901 · PMCID: PMC7299284
606. A Master Protocol Assessing the Safety, Tolerability, and Efficacy of Anti-Spike (S) SARS-CoV-2 Monoclonal Antibodies for the Treatment of Hospitalized Patients With COVID-19
Regeneron Pharmaceuticals clinicaltrials.gov (2021-02-03) https://clinicaltrials.gov/ct2/show/NCT04426695
607. A Master Protocol Assessing the Safety, Tolerability, and Efficacy of Anti-Spike (S) SARS-CoV-2 Monoclonal Antibodies for the Treatment of Ambulatory Patients With COVID-19
Regeneron Pharmaceuticals clinicaltrials.gov (2021-02-18) https://clinicaltrials.gov/ct2/show/NCT04425629
608. REGN-COV2, a Neutralizing Antibody Cocktail, in Outpatients with Covid-19
David M. Weinreich, Sumathi Sivapalasingam, Thomas Norton, Shazia Ali, Haitao Gao, Rafia Bhore, Bret J. Musser, Yuhwen Soo, Diana Rofail, Joseph Im, … George D. Yancopoulos New England Journal of Medicine (2021-01-21) https://doi.org/gh4sjh
DOI: 10.1056/nejmoa2035002 · PMID: 33332778 · PMCID: PMC7781102
609. Identification of SARS-CoV-2 spike mutations that attenuate monoclonal and serum antibody neutralization
Zhuoming Liu, Laura A. VanBlargan, Louis-Marie Bloyet, Paul W. Rothlauf, Rita E. Chen, Spencer Stumpf, Haiyan Zhao, John M. Errico, Elitza S. Theel, Mariel J. Liebeskind, … Sean P. J. Whelan Cell Host & Microbe (2021-03) https://doi.org/gh4m7j
DOI: 10.1016/j.chom.2021.01.014 · PMID: 33535027 · PMCID: PMC7839837
610. SARS-CoV-2 variants show resistance to neutralization by many monoclonal and serum-derived polyclonal antibodies
Michael Diamond, Rita Chen, Xuping Xie, James Case, Xianwen Zhang, Laura VanBlargan, Yang Liu, Jianying Liu, John Errico, Emma Winkler, … Pavlo Gilchuk Research Square (2021-02-09) https://doi.org/gh4sjz
DOI: 10.21203/rs.3.rs-228079/v1 · PMID: 33594356 · PMCID: PMC7885928
611. Impact of the B.1.1.7 variant on neutralizing monoclonal antibodies recognizing diverse epitopes on SARS–CoV–2 Spike
Carl Graham, Jeffrey Seow, Isabella Huettner, Hataf Khan, Neophytos Kouphou, Sam Acors, Helena Winstone, Suzanne Pickering, Rui Pedro Galao, Maria Jose Lista, … Katie J Doores Cold Spring Harbor Laboratory (2021-02-03) https://doi.org/gh4sjq
DOI: 10.1101/2021.02.03.429355 · PMID: 33564766 · PMCID: PMC7872354
612. Antibody Resistance of SARS-CoV-2 Variants B.1.351 and B.1.1.7
Pengfei Wang, Manoj S. Nair, Lihong Liu, Sho Iketani, Yang Luo, Yicheng Guo, Maple Wang, Jian Yu, Baoshan Zhang, Peter D. Kwong, … David D. Ho Cold Spring Harbor Laboratory (2021-02-12) https://doi.org/gh4sjp
DOI: 10.1101/2021.01.25.428137 · PMID: 33532778 · PMCID: PMC7852271
613. HIV-1 Broadly Neutralizing Antibody Extracts Its Epitope from a Kinked gp41 Ectodomain Region on the Viral Membrane
Zhen-Yu J. Sun, Kyoung Joon Oh, Mikyung Kim, Jessica Yu, Vladimir Brusic, Likai Song, Zhisong Qiao, Jia-huai Wang, Gerhard Wagner, Ellis L. Reinherz Immunity (2008-01) https://doi.org/ftw7t3
DOI: 10.1016/j.immuni.2007.11.018 · PMID: 18191596
614. Antibody Recognition of a Highly Conserved Influenza Virus Epitope
D. C. Ekiert, G. Bhabha, M.-A. Elsliger, R. H. E. Friesen, M. Jongeneelen, M. Throsby, J. Goudsmit, I. A. Wilson Science (2009-04-10) https://doi.org/ffsb4r
DOI: 10.1126/science.1171491 · PMID: 19251591 · PMCID: PMC2758658
615. Identification and characterization of novel neutralizing epitopes in the receptor-binding domain of SARS-CoV spike protein: Revealing the critical antigenic determinants in inactivated SARS-CoV vaccine
Yuxian He, Jingjing Li, Lanying Du, Xuxia Yan, Guangan Hu, Yusen Zhou, Shibo Jiang Vaccine (2006-06) https://doi.org/b99b68
DOI: 10.1016/j.vaccine.2006.04.054 · PMID: 16725238 · PMCID: PMC7115380
616. Broad and potent activity against SARS-like viruses by an engineered human monoclonal antibody
C. Garrett Rappazzo, Longping V. Tse, Chengzi I. Kaku, Daniel Wrapp, Mrunal Sakharkar, Deli Huang, Laura M. Deveau, Thomas J. Yockachonis, Andrew S. Herbert, Michael B. Battles, … Laura M. Walker Science (2021-02-19) https://doi.org/fsbc
DOI: 10.1126/science.abf4830 · PMID: 33495307 · PMCID: PMC7963221
617. The antiviral effect of interferon-beta against SARS-Coronavirus is not mediated by MxA protein
Martin Spiegel, Andreas Pichlmair, Elke Mühlberger, Otto Haller, Friedemann Weber Journal of Clinical Virology (2004-07) https://doi.org/cmc3ds
DOI: 10.1016/j.jcv.2003.11.013 · PMID: 15135736
618. Coronavirus virulence genes with main focus on SARS-CoV envelope gene
Marta L. DeDiego, Jose L. Nieto-Torres, Jose M. Jimenez-Guardeño, Jose A. Regla-Nava, Carlos Castaño-Rodriguez, Raul Fernandez-Delgado, Fernando Usera, Luis Enjuanes Virus Research (2014-12) https://doi.org/f6wm24
DOI: 10.1016/j.virusres.2014.07.024 · PMID: 25093995 · PMCID: PMC4261026
619. A Randomised Double-blind Placebo-controlled Trial to Determine the Safety and Efficacy of Inhaled SNG001 (IFN-β1a for Nebulisation) for the Treatment of Patients With Confirmed SARS-CoV-2 Infection
Synairgen Research Ltd. clinicaltrials.gov (2021-03-19) https://clinicaltrials.gov/ct2/show/NCT04385095
622. Safety and efficacy of inhaled nebulised interferon beta-1a (SNG001) for treatment of SARS-CoV-2 infection: a randomised, double-blind, placebo-controlled, phase 2 trial
Phillip D Monk, Richard J Marsden, Victoria J Tear, Jody Brookes, Toby N Batten, Marcin Mankowski, Felicity J Gabbay, Donna E Davies, Stephen T Holgate, Ling-Pei Ho, … Pedro MB Rodrigues The Lancet Respiratory Medicine (2021-02) https://doi.org/ghjzm4
DOI: 10.1016/s2213-2600(20)30511-7 · PMID: 33189161 · PMCID: PMC7836724
624. Effect of Intravenous Interferon β-1a on Death and Days Free From Mechanical Ventilation Among Patients With Moderate to Severe Acute Respiratory Distress Syndrome
V. Marco Ranieri, Ville Pettilä, Matti K. Karvonen, Juho Jalkanen, Peter Nightingale, David Brealey, Jordi Mancebo, Ricard Ferrer, Alain Mercat, Nicolò Patroniti, … for the INTEREST Study Group JAMA (2020-02-25) https://doi.org/ghzkww
DOI: 10.1001/jama.2019.22525 · PMID: 32065831
625. A Randomized Clinical Trial of the Efficacy and Safety of Interferon β-1a in Treatment of Severe COVID-19
Effat Davoudi-Monfared, Hamid Rahmani, Hossein Khalili, Mahboubeh Hajiabdolbaghi, Mohamadreza Salehi, Ladan Abbasian, Hossein Kazemzadeh, Mir Saeed Yekaninejad Antimicrobial Agents and Chemotherapy (2020-08-20) https://doi.org/gg5xvm
DOI: 10.1128/aac.01061-20 · PMID: 32661006 · PMCID: PMC7449227
626. A Multicenter, Adaptive, Randomized Blinded Controlled Trial of the Safety and Efficacy of Investigational Therapeutics for the Treatment of COVID-19 in Hospitalized Adults (ACTT-3)
National Institute of Allergy and Infectious Diseases (NIAID) clinicaltrials.gov (2021-02-04) https://clinicaltrials.gov/ct2/show/NCT04492475
627. COVID-19 PICU guidelines: for high- and limited-resource settings
Saraswati Kache, Mohammod Jobayer Chisti, Felicity Gumbo, Ezekiel Mupere, Xia Zhi, Karthi Nallasamy, Satoshi Nakagawa, Jan Hau Lee, Matteo Di Nardo, Pedro de la Oliva, … Joseph Carcillo Pediatric Research (2020-07-07) https://doi.org/gg4f3p
DOI: 10.1038/s41390-020-1053-9 · PMID: 32634818 · PMCID: PMC7577838
628. Table 1, Cost-Comparison Table for Biologic Disease-Modifying Drugs for Rheumatoid Arthritis
National Center for Biotechnology Information, U. S. National Library of Medicine 8600 Rockville Pike, Bethesda MD, 20894 Usa
(2015-08) https://www.ncbi.nlm.nih.gov/books/NBK349513/table/T43/
629. A Cost Comparison of Treatments of Moderate to Severe Psoriasis
Cheryl Hankin, Steven Feldman, Andy Szczotka, Randolph Stinger, Leslie Fish, David Hankin Drug Benefit Trends (2005-05) https://escholarship.umassmed.edu/meyers_pp/385
630. TNF-α inhibition for potential therapeutic modulation of SARS coronavirus infection
Edward Tobinick Current Medical Research and Opinion (2008-09-22) https://doi.org/bq4cx2
DOI: 10.1185/030079903125002757 · PMID: 14741070
633. COVID-19: combining antiviral and anti-inflammatory treatments
Justin Stebbing, Anne Phelan, Ivan Griffin, Catherine Tucker, Olly Oechsle, Dan Smith, Peter Richardson The Lancet Infectious Diseases (2020-04) https://doi.org/dph5
DOI: 10.1016/s1473-3099(20)30132-8 · PMID: 32113509 · PMCID: PMC7158903
634. Baricitinib as potential treatment for 2019-nCoV acute respiratory disease
Peter Richardson, Ivan Griffin, Catherine Tucker, Dan Smith, Olly Oechsle, Anne Phelan, Michael Rawling, Edward Savory, Justin Stebbing The Lancet (2020-02) https://doi.org/ggnrsx
DOI: 10.1016/s0140-6736(20)30304-4 · PMID: 32032529 · PMCID: PMC7137985
636. Baricitinib Combined With Antiviral Therapy in Symptomatic Patients Infected by COVID-19: an Open-label, Pilot Study
Fabrizio Cantini clinicaltrials.gov (2020-04-19) https://clinicaltrials.gov/ct2/show/NCT04320277
637. Design and Synthesis of Hydroxyferroquine Derivatives with Antimalarial and Antiviral Activities
Christophe Biot, Wassim Daher, Natascha Chavain, Thierry Fandeur, Jamal Khalife, Daniel Dive, Erik De Clercq Journal of Medicinal Chemistry (2006-05) https://doi.org/db4n83
DOI: 10.1021/jm0601856 · PMID: 16640347
638. An orally bioavailable broad-spectrum antiviral inhibits SARS-CoV-2 in human airway epithelial cell cultures and multiple coronaviruses in mice
Timothy P. Sheahan, Amy C. Sims, Shuntai Zhou, Rachel L. Graham, Andrea J. Pruijssers, Maria L. Agostini, Sarah R. Leist, Alexandra Schäfer, Kenneth H. Dinnon, Laura J. Stevens, … Ralph S. Baric Science Translational Medicine (2020-04-29) https://doi.org/ggrqd2
DOI: 10.1126/scitranslmed.abb5883 · PMID: 32253226 · PMCID: PMC7164393
639. Antiviral Monoclonal Antibodies: Can They Be More Than Simple Neutralizing Agents?
Mireia Pelegrin, Mar Naranjo-Gomez, Marc Piechaczyk Trends in Microbiology (2015-10) https://doi.org/f7vzrf
DOI: 10.1016/j.tim.2015.07.005 · PMID: 26433697 · PMCID: PMC7127033
640. Intranasal Treatment with Poly(I{middle dot}C) Protects Aged Mice from Lethal Respiratory Virus Infections
J. Zhao, C. Wohlford-Lenane, J. Zhao, E. Fleming, T. E. Lane, P. B. McCray, S. Perlman Journal of Virology (2012-08-22) https://doi.org/f4bzfp
DOI: 10.1128/jvi.01410-12 · PMID: 22915814 · PMCID: PMC3486278
642. COVID-19: The Inflammation Link and the Role of Nutrition in Potential Mitigation
Ioannis Zabetakis, Ronan Lordan, Catherine Norton, Alexandros Tsoupras Nutrients (2020-05-19) https://doi.org/ggxdq3
DOI: 10.3390/nu12051466 · PMID: 32438620 · PMCID: PMC7284818
643. Could nutrition modulate COVID-19 susceptibility and severity of disease? A systematic review
Philip T. James, Zakari Ali, Andrew E. Armitage, Ana Bonell, Carla Cerami, Hal Drakesmith, Modou Jobe, Kerry S. Jones, Zara Liew, Sophie E. Moore, … Andrew M. Prentice Cold Spring Harbor Laboratory (2020-10-21) https://doi.org/ghr94g
DOI: 10.1101/2020.10.19.20214395
645. Nutritional status of patients with COVID-19
Jae Hyoung Im, Young Soo Je, Jihyeon Baek, Moon-Hyun Chung, Hea Yoon Kwon, Jin-Soo Lee International Journal of Infectious Diseases (2020-11) https://doi.org/gg7t5t
DOI: 10.1016/j.ijid.2020.08.018 · PMID: 32795605 · PMCID: PMC7418699
646. Optimal Nutritional Status for a Well-Functioning Immune System Is an Important Factor to Protect against Viral Infections
Philip Calder, Anitra Carr, Adrian Gombart, Manfred Eggersdorfer Nutrients (2020-04-23) https://doi.org/gg29hh
DOI: 10.3390/nu12041181 · PMID: 32340216 · PMCID: PMC7230749
654. Structural Design Principles for Delivery of Bioactive Components in Nutraceuticals and Functional Foods
David Julian McClements, Eric Andrew Decker, Yeonhwa Park, Jochen Weiss Critical Reviews in Food Science and Nutrition (2009-06-16) https://doi.org/dt68m4
DOI: 10.1080/10408390902841529 · PMID: 19484636
659. Nutraceuticals - shedding light on the grey area between pharmaceuticals and food
Antonello Santini, Ettore Novellino Expert Review of Clinical Pharmacology (2018-04-23) https://doi.org/ggwztk
DOI: 10.1080/17512433.2018.1464911 · PMID: 29667442
663. Nutraceuticals: opening the debate for a regulatory framework
Antonello Santini, Silvia Miriam Cammarata, Giacomo Capone, Angela Ianaro, Gian Carlo Tenore, Luca Pani, Ettore Novellino British Journal of Clinical Pharmacology (2018-04) https://doi.org/ggwztm
DOI: 10.1111/bcp.13496 · PMID: 29433155 · PMCID: PMC5867125
664. Reviewing the Nutrition and Health Claims Regulation (EC) No. 1924/2006: What do we know about its challenges and potential impact on innovation?
Stefanie Bröring, Sukhada Khedkar, Stefano Ciliberti International Journal of Food Sciences and Nutrition (2016-08-02) https://doi.org/ghr936
DOI: 10.1080/09637486.2016.1212816 · PMID: 27484163
671. Reducing mortality from 2019-nCoV: host-directed therapies should be an option
Alimuddin Zumla, David S Hui, Esam I Azhar, Ziad A Memish, Markus Maeurer The Lancet (2020-02) https://doi.org/ggkd3b
DOI: 10.1016/s0140-6736(20)30305-6
672. Diet Supplementation, Probiotics, and Nutraceuticals in SARS-CoV-2 Infection: A Scoping Review
Fabio Infusino, Massimiliano Marazzato, Massimo Mancone, Francesco Fedele, Claudio Maria Mastroianni, Paolo Severino, Giancarlo Ceccarelli, Letizia Santinelli, Elena Cavarretta, Antonino G. M. Marullo, … Gabriella d’Ettorre Nutrients (2020-06-08) https://doi.org/gg8k58
DOI: 10.3390/nu12061718 · PMID: 32521760 · PMCID: PMC7352781
673. Potential interventions for novel coronavirus in China: A systematic review
Lei Zhang, Yunhui Liu Journal of Medical Virology (2020-03-03) https://doi.org/ggpx57
DOI: 10.1002/jmv.25707 · PMID: 32052466
674. Nutraceuticals have potential for boosting the type 1 interferon response to RNA viruses including influenza and coronavirus
Mark F. McCarty, James J. DiNicolantonio Progress in Cardiovascular Diseases (2020-05) https://doi.org/ggpwx2
DOI: 10.1016/j.pcad.2020.02.007 · PMID: 32061635
675. Inflammation and cardiovascular disease: are marine phospholipids the answer?
Ronan Lordan, Shane Redfern, Alexandros Tsoupras, Ioannis Zabetakis Food & Function (2020) https://doi.org/gg29hg
DOI: 10.1039/c9fo01742a · PMID: 32270798
676. The Potential Beneficial Effect of EPA and DHA Supplementation Managing Cytokine Storm in Coronavirus Disease
Zoltán Szabó, Tamás Marosvölgyi, Éva Szabó, Péter Bai, Mária Figler, Zsófia Verzár Frontiers in Physiology (2020-06-19) https://doi.org/gg4hz4
DOI: 10.3389/fphys.2020.00752 · PMID: 32636763 · PMCID: PMC7318894
677. Exploitation of Microalgae Species for Nutraceutical Purposes: Cultivation Aspects
Sushanta Saha, Patrick Murray Fermentation (2018-06-14) https://doi.org/ghv64j
DOI: 10.3390/fermentation4020046
678. Prospective options of algae-derived nutraceuticals as supplements to combat COVID-19 and human coronavirus diseases
Sachitra K. Ratha, Nirmal Renuka, Ismail Rawat, Faizal Bux Nutrition (2021-03) https://doi.org/ghr93z
DOI: 10.1016/j.nut.2020.111089 · PMID: 33412367 · PMCID: PMC7680017
679. Safety Aspects of Fish Oils
Erik Berg Schmidt, Jørn Munkhof Møller, Niels Svaneborg, Jørn Dyerberg Drug Investigation (2012-10-14) https://doi.org/ghvqm8
DOI: 10.1007/bf03257413
683. Marine omega-3 fatty acids and inflammatory processes: Effects, mechanisms and clinical relevance
Philip C. Calder Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids (2015-04) https://doi.org/gf8pc6
DOI: 10.1016/j.bbalip.2014.08.010 · PMID: 25149823
684. N-3 polyunsaturated fatty acids modulate B cell activity in pre-clinical models: Implications for the immune response to infections
Jarrett Whelan, Kymberly M. Gowdy, Saame Raza Shaikh European Journal of Pharmacology (2016-08) https://doi.org/f8xn7q
DOI: 10.1016/j.ejphar.2015.03.100 · PMID: 26022530 · PMCID: PMC4662641
685. Blood omega-3 fatty acids and death from COVID-19: A pilot study
Arash Asher, Nathan L. Tintle, Michael Myers, Laura Lockshon, Heribert Bacareza, William S. Harris Prostaglandins, Leukotrienes and Essential Fatty Acids (2021-03) https://doi.org/ghv63m
DOI: 10.1016/j.plefa.2021.102250 · PMID: 33516093 · PMCID: PMC7816864
686. n-3 Polyunsaturated Fatty Acids Improve Inflammation via Inhibiting Sphingosine Kinase 1 in a Rat Model of Parenteral Nutrition and CLP-Induced Sepsis
Tao Tian, Yunzhao Zhao, Qian Huang, Jieshou Li Lipids (2016-02-08) https://doi.org/ghvqm9
DOI: 10.1007/s11745-016-4129-x · PMID: 26856322
688. Clinical characteristics of 82 cases of death from COVID-19
Bicheng Zhang, Xiaoyang Zhou, Yanru Qiu, Yuxiao Song, Fan Feng, Jia Feng, Qibin Song, Qingzhu Jia, Jun Wang PLOS ONE (2020-07-09) https://doi.org/gg4sgx
DOI: 10.1371/journal.pone.0235458 · PMID: 32645044 · PMCID: PMC7347130
689. Effects of an omega-3 fatty acid-enriched lipid emulsion on eicosanoid synthesis in acute respiratory distress syndrome (ARDS): A prospective, randomized, double-blind, parallel group study
Joan Sabater, Joan Masclans, Judit Sacanell, Pilar Chacon, Pilar Sabin, Mercè Planas Nutrition & Metabolism (2011) https://doi.org/bg2kpp
DOI: 10.1186/1743-7075-8-22 · PMID: 21477318 · PMCID: PMC3080285
690. Immunonutrition for Adults With ARDS: Results From a Cochrane Systematic Review and Meta-Analysis
Ahilanandan Dushianthan, Rebecca Cusack, Victoria A Burgess, Michael PW Grocott, Philip Calder Respiratory Care (2020-01) https://doi.org/fj2p
DOI: 10.4187/respcare.06965 · PMID: 31506339
691. Correlation analysis of omega-3 fatty acids and mortality of sepsis and sepsis-induced ARDS in adults: data from previous randomized controlled trials
HuaiSheng Chen, Su Wang, Ying Zhao, YuTian Luo, HuaSheng Tong, Lei Su Nutrition Journal (2018-05-31) https://doi.org/gdpjtq
DOI: 10.1186/s12937-018-0356-8 · PMID: 29859104 · PMCID: PMC5984323
693. Specialized pro-resolving mediators: endogenous regulators of infection and inflammation
Maria C. Basil, Bruce D. Levy Nature Reviews Immunology (2015-12-21) https://doi.org/f9fgtd
DOI: 10.1038/nri.2015.4 · PMID: 26688348 · PMCID: PMC5242505
696. The Specialized Proresolving Mediator 17-HDHA Enhances the Antibody-Mediated Immune Response against Influenza Virus: A New Class of Adjuvant?
Sesquile Ramon, Steven F. Baker, Julie M. Sahler, Nina Kim, Eric A. Feldsott, Charles N. Serhan, Luis Martínez-Sobrido, David J. Topham, Richard P. Phipps The Journal of Immunology (2014-12-15) https://doi.org/f6spr8
DOI: 10.4049/jimmunol.1302795 · PMID: 25392529 · PMCID: PMC4258475
697. The Lipid Mediator Protectin D1 Inhibits Influenza Virus Replication and Improves Severe Influenza
Masayuki Morita, Keiji Kuba, Akihiko Ichikawa, Mizuho Nakayama, Jun Katahira, Ryo Iwamoto, Tokiko Watanebe, Saori Sakabe, Tomo Daidoji, Shota Nakamura, … Yumiko Imai Cell (2013-03) https://doi.org/f4rbgb
DOI: 10.1016/j.cell.2013.02.027 · PMID: 23477864
698. Inflammation resolution: a dual-pronged approach to averting cytokine storms in COVID-19?
Dipak Panigrahy, Molly M. Gilligan, Sui Huang, Allison Gartung, Irene Cortés-Puch, Patricia J. Sime, Richard P. Phipps, Charles N. Serhan, Bruce D. Hammock Cancer and Metastasis Reviews (2020-05-08) https://doi.org/ggvv7w
DOI: 10.1007/s10555-020-09889-4 · PMID: 32385712 · PMCID: PMC7207990
699. Pro resolving inflammatory effects of the lipid mediators of omega 3 fatty acids and its implication in SARS COVID-19
Pedro-Antonio Regidor, Fernando Gonzalez Santos, Jose Miguel Rizo, Fernando Moreno Egea Medical Hypotheses (2020-12) https://doi.org/ghr93x
DOI: 10.1016/j.mehy.2020.110340 · PMID: 33069094 · PMCID: PMC7543931
700. Obesity-Driven Deficiencies of Specialized Pro-resolving Mediators May Drive Adverse Outcomes During SARS-CoV-2 Infection
Anandita Pal, Kymberly M. Gowdy, Kenneth J. Oestreich, Melinda Beck, Saame Raza Shaikh Frontiers in Immunology (2020-08-11) https://doi.org/ght38j
DOI: 10.3389/fimmu.2020.01997 · PMID: 32983141 · PMCID: PMC7438933
701. Fish Oil-Fed Mice Have Impaired Resistance to Influenza Infection
Nicole M. J. Schwerbrock, Erik A. Karlsson, Qing Shi, Patricia A. Sheridan, Melinda A. Beck The Journal of Nutrition (2009-08) https://doi.org/dv45f4
DOI: 10.3945/jn.109.108027 · PMID: 19549756 · PMCID: PMC2709305
702. Modulation of host defence against bacterial and viral infections by omega-3 polyunsaturated fatty acids
Marie-Odile Husson, Delphine Ley, Céline Portal, Madeleine Gottrand, Thomas Hueso, Jean-Luc Desseyn, Frédéric Gottrand Journal of Infection (2016-12) https://doi.org/f9pp2h
DOI: 10.1016/j.jinf.2016.10.001 · PMID: 27746159
703. Bioactive products formed in humans from fish oils
Carsten Skarke, Naji Alamuddin, John A. Lawson, Xuanwen Li, Jane F. Ferguson, Muredach P. Reilly, Garret A. FitzGerald Journal of Lipid Research (2015-09) https://doi.org/f7pm5g
DOI: 10.1194/jlr.m060392 · PMID: 26180051 · PMCID: PMC4548785
704. Resolving Inflammatory Storm in COVID-19 Patients by Omega-3 Polyunsaturated Fatty Acids - A Single-blind, Randomized, Placebo-controlled Feasibility Study
Magnus Bäck clinicaltrials.gov (2020-11-27) https://clinicaltrials.gov/ct2/show/NCT04647604
705. Stimulating the Resolution of Inflammation Through Omega-3 Polyunsaturated Fatty Acids in COVID-19: Rationale for the COVID-Omega-F Trial
Hildur Arnardottir, Sven-Christian Pawelzik, Ulf Öhlund Wistbacka, Gonzalo Artiach, Robin Hofmann, Ingalill Reinholdsson, Frieder Braunschweig, Per Tornvall, Dorota Religa, Magnus Bäck Frontiers in Physiology (2021-01-11) https://doi.org/ghv64h
DOI: 10.3389/fphys.2020.624657 · PMID: 33505321 · PMCID: PMC7830247
707. COVID-19 and Thrombotic or Thromboembolic Disease: Implications for Prevention, Antithrombotic Therapy, and Follow-Up
Behnood Bikdeli, Mahesh V. Madhavan, David Jimenez, Taylor Chuich, Isaac Dreyfus, Elissa Driggin, Caroline Der Nigoghossian, Walter Ageno, Mohammad Madjid, Yutao Guo, … Gregory Y. H. Lip Journal of the American College of Cardiology (2020-06) https://doi.org/ggsppk
DOI: 10.1016/j.jacc.2020.04.031 · PMID: 32311448 · PMCID: PMC7164881
710. Platelet activation and prothrombotic mediators at the nexus of inflammation and atherosclerosis: Potential role of antiplatelet agents
Ronan Lordan, Alexandros Tsoupras, Ioannis Zabetakis Blood Reviews (2020-04) https://doi.org/ggvv7x
DOI: 10.1016/j.blre.2020.100694 · PMID: 32340775
711. An Investigation on the Effects of Icosapent Ethyl (VascepaTM) on Inflammatory Biomarkers in Individuals With COVID-19 - Full Text View - ClinicalTrials.govhttps://clinicaltrials.gov/ct2/show/NCT04412018
712. Cardiovascular Risk Reduction with Icosapent Ethyl for Hypertriglyceridemia
Deepak L. Bhatt, P. Gabriel Steg, Michael Miller, Eliot A. Brinton, Terry A. Jacobson, Steven B. Ketchum, Ralph T. Doyle, Rebecca A. Juliano, Lixia Jiao, Craig Granowitz, … Christie M. Ballantyne New England Journal of Medicine (2019-01-03) https://doi.org/gfj3w9
DOI: 10.1056/nejmoa1812792 · PMID: 30415628
713. A Randomised, Double-blind, Placebo Controlled Study of Eicosapentaenoic Acid (EPA-FFA) Gastro-resistant Capsules to Treat Hospitalised Subjects With Confirmed SARS-CoV-2
S.L.A. Pharma AG clinicaltrials.gov (2020-10-29) https://clinicaltrials.gov/ct2/show/NCT04335032
714. Anti-inflammatory/Antioxidant Oral Nutrition Supplementation on the Cytokine Storm and Progression of COVID-19: A Randomized Controlled Trial
Mahmoud Abulmeaty FACN M. D. clinicaltrials.gov (2020-09-18) https://clinicaltrials.gov/ct2/show/NCT04323228
715. Functional Role of Dietary Intervention to Improve the Outcome of COVID-19: A Hypothesis of Work
Giovanni Messina, Rita Polito, Vincenzo Monda, Luigi Cipolloni, Nunzio Di Nunno, Giulio Di Mizio, Paolo Murabito, Marco Carotenuto, Antonietta Messina, Daniela Pisanelli, … Francesco Sessa International Journal of Molecular Sciences (2020-04-28) https://doi.org/ggvb88
DOI: 10.3390/ijms21093104 · PMID: 32354030 · PMCID: PMC7247152
717. Zinc-Dependent Suppression of TNF-α Production Is Mediated by Protein Kinase A-Induced Inhibition of Raf-1, IκB Kinase β, and NF-κB
Verena von Bülow, Svenja Dubben, Gabriela Engelhardt, Silke Hebel, Birgit Plümäkers, Holger Heine, Lothar Rink, Hajo Haase The Journal of Immunology (2007-09-15) https://doi.org/f3vs45
DOI: 10.4049/jimmunol.179.6.4180 · PMID: 17785857
718. Zinc activates NF-κB in HUT-78 cells
Ananda S. Prasad, Bin Bao, Frances W. J. Beck, Fazlul H. Sarkar Journal of Laboratory and Clinical Medicine (2001-10) https://doi.org/cnc6fr
DOI: 10.1067/mlc.2001.118108 · PMID: 11574819
719. Innate or Adaptive Immunity? The Example of Natural Killer Cells
E. Vivier, D. H. Raulet, A. Moretta, M. A. Caligiuri, L. Zitvogel, L. L. Lanier, W. M. Yokoyama, S. Ugolini Science (2011-01-06) https://doi.org/ckzg9g
DOI: 10.1126/science.1198687 · PMID: 21212348 · PMCID: PMC3089969
720. Zinc supplementation decreases incidence of infections in the elderly: effect of zinc on generation of cytokines and oxidative stress
Ananda S Prasad, Frances WJ Beck, Bin Bao, James T Fitzgerald, Diane C Snell, Joel D Steinberg, Lavoisier J Cardozo The American Journal of Clinical Nutrition (2007-03) https://doi.org/ggqmgs
DOI: 10.1093/ajcn/85.3.837 · PMID: 17344507
725. Zn2+ Inhibits Coronavirus and Arterivirus RNA Polymerase Activity In Vitro and Zinc Ionophores Block the Replication of These Viruses in Cell Culture
Aartjan J. W. te Velthuis, Sjoerd H. E. van den Worm, Amy C. Sims, Ralph S. Baric, Eric J. Snijder, Martijn J. van Hemert PLoS Pathogens (2010-11-04) https://doi.org/d95x4g
DOI: 10.1371/journal.ppat.1001176 · PMID: 21079686 · PMCID: PMC2973827
726. The SARS-coronavirus papain-like protease: Structure, function and inhibition by designed antiviral compounds
Yahira M. Báez-Santos, Sarah E. St. John, Andrew D. Mesecar Antiviral Research (2015-03) https://doi.org/f63hjp
DOI: 10.1016/j.antiviral.2014.12.015 · PMID: 25554382 · PMCID: PMC5896749
727. A Randomized Study Evaluating the Safety and Efficacy of Hydroxychloroquine and Zinc in Combination With Either Azithromycin or Doxycycline for the Treatment of COVID-19 in the Outpatient Setting
Avni Thakore MD clinicaltrials.gov (2020-12-08) https://clinicaltrials.gov/ct2/show/NCT04370782
728. A Study of Hydroxychloroquine and Zinc in the Prevention of COVID-19 Infection in Military Healthcare Workers - Full Text View - ClinicalTrials.govhttps://clinicaltrials.gov/ct2/show/NCT04377646
729. Therapies to Prevent Progression of COVID-19, Including Hydroxychloroquine, Azithromycin, Zinc, Vitamin D, Vitamin B12 With or Without Vitamin C, a Multi-centre, International, Randomized Trial: The International ALLIANCE Study
National Institute of Integrative Medicine, Australia clinicaltrials.gov (2020-09-09) https://clinicaltrials.gov/ct2/show/NCT04395768
731. Hydroxychloroquine with or without Azithromycin in Mild-to-Moderate Covid-19
Alexandre B. Cavalcanti, Fernando G. Zampieri, Regis G. Rosa, Luciano C. P. Azevedo, Viviane C. Veiga, Alvaro Avezum, Lucas P. Damiani, Aline Marcadenti, Letícia Kawano-Dourado, Thiago Lisboa, … Otavio Berwanger New England Journal of Medicine (2020-11-19) https://doi.org/gg5343
DOI: 10.1056/nejmoa2019014 · PMID: 32706953 · PMCID: PMC7397242
732. The efficacy and safety of hydroxychloroquine for COVID-19 prophylaxis: A systematic review and meta-analysis of randomized trials
Kimberley Lewis, Dipayan Chaudhuri, Fayez Alshamsi, Laiya Carayannopoulos, Karin Dearness, Zain Chagla, Waleed Alhazzani, for the GUIDE Group PLOS ONE (2021-01-06) https://doi.org/ghsv36
DOI: 10.1371/journal.pone.0244778 · PMID: 33406138 · PMCID: PMC7787432
733. Effect of hydroxychloroquine with or without azithromycin on the mortality of coronavirus disease 2019 (COVID-19) patients: a systematic review and meta-analysis
Thibault Fiolet, Anthony Guihur, Mathieu Edouard Rebeaud, Matthieu Mulot, Nathan Peiffer-Smadja, Yahya Mahamat-Saleh Clinical Microbiology and Infection (2021-01) https://doi.org/gg9jk2
DOI: 10.1016/j.cmi.2020.08.022 · PMID: 32860962 · PMCID: PMC7449662
734. Zinc sulfate in combination with a zinc ionophore may improve outcomes in hospitalized COVID-19 patients
Philip M. Carlucci, Tania Ahuja, Christopher Petrilli, Harish Rajagopalan, Simon Jones, Joseph Rahimian Journal of Medical Microbiology (2020-10-01) https://doi.org/ghnws7
DOI: 10.1099/jmm.0.001250 · PMID: 32930657 · PMCID: PMC7660893
735. The Minimal Effect of Zinc on the Survival of Hospitalized Patients With COVID-19
Jasper Seth Yao, Joseph Alexander Paguio, Edward Christopher Dee, Hanna Clementine Tan, Achintya Moulick, Carmelo Milazzo, Jerry Jurado, Nicolás Della Penna, Leo Anthony Celi Chest (2021-01) https://doi.org/gg5w36
DOI: 10.1016/j.chest.2020.06.082 · PMID: 32710890 · PMCID: PMC7375307
736. Coronavirus Disease 2019- Using Ascorbic Acid and Zinc Supplementation (COVIDAtoZ) Research Study A Randomized, Open Label Single Center Study
Milind Desai clinicaltrials.gov (2021-01-28) https://clinicaltrials.gov/ct2/show/NCT04342728
737. Vitamin B12 May Inhibit RNA-Dependent-RNA Polymerase Activity of nsp12 from the COVID-19 Virus
Naveen Narayanan, Deepak T. Nair Preprints (2020-03-22) https://doi.org/ggqmjc
DOI: 10.20944/preprints202003.0347.v1
738. The Long History of Vitamin C: From Prevention of the Common Cold to Potential Aid in the Treatment of COVID-19
Giuseppe Cerullo, Massimo Negro, Mauro Parimbelli, Michela Pecoraro, Simone Perna, Giorgio Liguori, Mariangela Rondanelli, Hellas Cena, Giuseppe D’Antona Frontiers in Immunology (2020-10-28) https://doi.org/ghr943
DOI: 10.3389/fimmu.2020.574029 · PMID: 33193359 · PMCID: PMC7655735
740. Vitamin C Mitigates Oxidative Stress and Tumor Necrosis Factor-Alpha in Severe Community-Acquired Pneumonia and LPS-Induced Macrophages
Yuanyuan Chen, Guangyan Luo, Jiao Yuan, Yuanyuan Wang, Xiaoqiong Yang, Xiaoyun Wang, Guoping Li, Zhiguang Liu, Nanshan Zhong Mediators of Inflammation (2014) https://doi.org/f6nb5f
DOI: 10.1155/2014/426740 · PMID: 25253919 · PMCID: PMC4165740
741. Intravenous infusion of ascorbic acid decreases serum histamine concentrations in patients with allergic and non-allergic diseases
Alexander F. Hagel, Christian M. Layritz, Wolfgang H. Hagel, Hans-Jürgen Hagel, Edith Hagel, Wolfgang Dauth, Jürgen Kressel, Tanja Regnet, Andreas Rosenberg, Markus F. Neurath, … Martin Raithel Naunyn-Schmiedeberg’s Archives of Pharmacology (2013-05-11) https://doi.org/f48jsb
DOI: 10.1007/s00210-013-0880-1 · PMID: 23666445
745. Metabolism of ascorbic acid (vitamin C) in subjects infected with common cold viruses
J. E. W. Davies, R. E. Hughes, Eleri Jones, Sylvia E. Reed, J. W. Craig, D. A. J. Tyrrell Biochemical Medicine (1979-02) https://doi.org/fd22sv
DOI: 10.1016/0006-2944(79)90058-9
749. Serum Levels of Vitamin C and Vitamin D in a Cohort of Critically Ill COVID-19 Patients of a North American Community Hospital Intensive Care Unit in May 2020: A Pilot Study
Cristian Arvinte, Maharaj Singh, Paul E. Marik Medicine in Drug Discovery (2020-12) https://doi.org/ghnwqt
DOI: 10.1016/j.medidd.2020.100064 · PMID: 32964205 · PMCID: PMC7499070
750. Vitamin C levels in patients with SARS-CoV-2-associated acute respiratory distress syndrome
Luis Chiscano-Camón, Juan Carlos Ruiz-Rodriguez, Adolf Ruiz-Sanmartin, Oriol Roca, Ricard Ferrer Critical Care (2020-08-26) https://doi.org/ghbr97
DOI: 10.1186/s13054-020-03249-y · PMID: 32847620 · PMCID: PMC7447967
751. Targeting coagulation activation in severe COVID-19 pneumonia: lessons from bacterial pneumonia and sepsis
Ricardo J. José, Andrew Williams, Ari Manuel, Jeremy S. Brown, Rachel C. Chambers European Respiratory Review (2020-10-01) https://doi.org/ghr94s
DOI: 10.1183/16000617.0240-2020 · PMID: 33004529 · PMCID: PMC7537941
753. The use of IV vitamin C for patients with COVID-19: a case series
Raul Hiedra, Kevin Bryan Lo, Mohammad Elbashabsheh, Fahad Gul, Robert Matthew Wright, Jeri Albano, Zurab Azmaiparashvili, Gabriel Patarroyo Aponte Expert Review of Anti-infective Therapy (2020-08-01) https://doi.org/ghr938
DOI: 10.1080/14787210.2020.1794819 · PMID: 32662690 · PMCID: PMC7441798
756. Effect of Vitamin C Infusion on Organ Failure and Biomarkers of Inflammation and Vascular Injury in Patients With Sepsis and Severe Acute Respiratory Failure
Alpha A. Fowler, Jonathon D. Truwit, R. Duncan Hite, Peter E. Morris, Christine DeWilde, Anna Priday, Bernard Fisher, Leroy R. Thacker, Ramesh Natarajan, Donald F. Brophy, … Matthew Halquist JAMA (2019-10-01) https://doi.org/ggqmh8
DOI: 10.1001/jama.2019.11825 · PMID: 31573637 · PMCID: PMC6777268
757. Intravenous high-dose vitamin C for the treatment of severe COVID-19: study protocol for a multicentre randomised controlled trial
Fang Liu, Yuan Zhu, Jing Zhang, Yiming Li, Zhiyong Peng BMJ Open (2020-07-08) https://doi.org/gg4sgj
DOI: 10.1136/bmjopen-2020-039519 · PMID: 32641343 · PMCID: PMC7348463
758. Pilot Trial of High-dose vitamin C in critically ill COVID-19 patients
Jing Zhang, Xin Rao, Yiming Li, Yuan Zhu, Fang Liu, Guangling Guo, Guoshi Luo, Zhongji Meng, Daniel De Backer, Hui Xiang, Zhi-Yong Peng Research Square (2020-08-03) https://doi.org/ghr94x
DOI: 10.21203/rs.3.rs-52778/v2
759. High-dose vitamin C infusion for the treatment of critically ill COVID-19
Jing Zhang, Xin Rao, Yiming Li, Yuan Zhu, Fang Liu, Guangling Guo, Guoshi Luo, Zhongji Meng, Daniel De Backer, Hui Xiang, Zhi-Yong Peng Research Square (2020-08-03) https://doi.org/ghr94v
DOI: 10.21203/rs.3.rs-52778/v1
760. Dietary Reference Intakes for Vitamin C, Vitamin E, Selenium, and Carotenoids
Panel on Dietary Antioxidants and Related Compounds, Subcommittee on Upper Reference Levels of Nutrients, Subcommittee on Interpretation and Uses of Dietary Reference Intakes, Standing Committee on the Scientific Evaluation of Dietary Reference Intakes, Food and Nutrition Board, Institute of Medicine The National Academies Press (2000-07-27) https://doi.org/ghtvqx
DOI: 10.17226/9810 · PMID: 25077263
761. Vitamin D and Infectious Diseases: Simple Bystander or Contributing Factor?
Pedro Gois, Daniela Ferreira, Simon Olenski, Antonio Seguro Nutrients (2017-06-24) https://doi.org/ggpcwr
DOI: 10.3390/nu9070651 · PMID: 28672783 · PMCID: PMC5537771
765. Regulation of Immune Function by Vitamin D and Its Use in Diseases of Immunity
An-Sofie Vanherwegen, Conny Gysemans, Chantal Mathieu Endocrinology and Metabolism Clinics of North America (2017-12) https://doi.org/gcm7h9
DOI: 10.1016/j.ecl.2017.07.010 · PMID: 29080635
767. Vitamin D in the prevention of acute respiratory infection: Systematic review of clinical studies
David A. Jolliffe, Christopher J. Griffiths, Adrian R. Martineau The Journal of Steroid Biochemistry and Molecular Biology (2013-07) https://doi.org/ggqmh9
DOI: 10.1016/j.jsbmb.2012.11.017 · PMID: 23220552
768. Vitamin D: modulator of the immune system
Femke Baeke, Tatiana Takiishi, Hannelie Korf, Conny Gysemans, Chantal Mathieu Current Opinion in Pharmacology (2010-08) https://doi.org/d43qtf
DOI: 10.1016/j.coph.2010.04.001 · PMID: 20427238
769. Evidence that Vitamin D Supplementation Could Reduce Risk of Influenza and COVID-19 Infections and Deaths
William B. Grant, Henry Lahore, Sharon L. McDonnell, Carole A. Baggerly, Christine B. French, Jennifer L. Aliano, Harjit P. Bhattoa Nutrients (2020-04-02) https://doi.org/ggr2v5
DOI: 10.3390/nu12040988 · PMID: 32252338 · PMCID: PMC7231123
770. Perspective: Vitamin D deficiency and COVID‐19 severity – plausibly linked by latitude, ethnicity, impacts on cytokines, ACE2 and thrombosis
J. M. Rhodes, S. Subramanian, E. Laird, G. Griffin, R. A. Kenny Journal of Internal Medicine (2020-07-22) https://doi.org/ghc7dh
DOI: 10.1111/joim.13149 · PMID: 32613681 · PMCID: PMC7361294
772. Editorial: low population mortality from COVID-19 in countries south of latitude 35 degrees North supports vitamin D as a factor determining severity
Jonathan M. Rhodes, Sreedhar Subramanian, Eamon Laird, Rose A. Kenny Alimentary Pharmacology & Therapeutics (2020-06) https://doi.org/ggtw4b
DOI: 10.1111/apt.15777 · PMID: 32311755 · PMCID: PMC7264531
773. 25-Hydroxyvitamin D Concentrations Are Lower in Patients with Positive PCR for SARS-CoV-2
Antonio D’Avolio, Valeria Avataneo, Alessandra Manca, Jessica Cusato, Amedeo De Nicolò, Renzo Lucchini, Franco Keller, Marco Cantù Nutrients (2020-05-09) https://doi.org/ggvv76
DOI: 10.3390/nu12051359 · PMID: 32397511 · PMCID: PMC7285131
774. Vitamin D deficiency as risk factor for severe COVID-19: a convergence of two pandemics
D. De Smet, K. De Smet, P. Herroelen, S. Gryspeerdt, G. A. Martens Cold Spring Harbor Laboratory (2020-05-05) https://doi.org/ggvv75
DOI: 10.1101/2020.05.01.20079376
775. Vitamin D sufficiency, a serum 25-hydroxyvitamin D at least 30 ng/mL reduced risk for adverse clinical outcomes in patients with COVID-19 infection
Zhila Maghbooli, Mohammad Ali Sahraian, Mehdi Ebrahimi, Marzieh Pazoki, Samira Kafan, Hedieh Moradi Tabriz, Azar Hadadi, Mahnaz Montazeri, Mehrad Nasiri, Arash Shirvani, Michael F. Holick PLOS ONE (2020-09-25) https://doi.org/ghdzx8
DOI: 10.1371/journal.pone.0239799 · PMID: 32976513 · PMCID: PMC7518605
777. Low plasma 25(OH) vitamin D level is associated with increased risk of COVID‐19 infection: an Israeli population‐based study
Eugene Merzon, Dmitry Tworowski, Alessandro Gorohovski, Shlomo Vinker, Avivit Golan Cohen, Ilan Green, Milana Frenkel‐Morgenstern The FEBS Journal (2020-08-28) https://doi.org/gg7b5c
DOI: 10.1111/febs.15495 · PMID: 32700398 · PMCID: PMC7404739
778. Association of Vitamin D Status and Other Clinical Characteristics With COVID-19 Test Results
David O. Meltzer, Thomas J. Best, Hui Zhang, Tamara Vokes, Vineet Arora, Julian Solway JAMA Network Open (2020-09-03) https://doi.org/ghdzw6
DOI: 10.1001/jamanetworkopen.2020.19722 · PMID: 32880651 · PMCID: PMC7489852
779. Vitamin D Status in Hospitalized Patients with SARS-CoV-2 Infection
José L Hernández, Daniel Nan, Marta Fernandez-Ayala, Mayte García-Unzueta, Miguel A Hernández-Hernández, Marcos López-Hoyos, Pedro Muñoz-Cacho, José M Olmos, Manuel Gutiérrez-Cuadra, Juan J Ruiz-Cubillán, … Víctor M Martínez-Taboada The Journal of Clinical Endocrinology & Metabolism (2020-10-27) https://doi.org/ghh737
DOI: 10.1210/clinem/dgaa733 · PMID: 33159440 · PMCID: PMC7797757
780. Analysis of vitamin D level among asymptomatic and critically ill COVID-19 patients and its correlation with inflammatory markers
Anshul Jain, Rachna Chaurasia, Narendra Singh Sengar, Mayank Singh, Sachin Mahor, Sumit Narain Scientific Reports (2020-11-19) https://doi.org/ghm3zn
DOI: 10.1038/s41598-020-77093-z · PMID: 33214648 · PMCID: PMC7677378
781. Low 25-Hydroxyvitamin D Levels on Admission to the Intensive Care Unit May Predispose COVID-19 Pneumonia Patients to a Higher 28-Day Mortality Risk: A Pilot Study on a Greek ICU Cohort
Alice G. Vassiliou, Edison Jahaj, Maria Pratikaki, Stylianos E. Orfanos, Ioanna Dimopoulou, Anastasia Kotanidou Nutrients (2020-12-09) https://doi.org/ghr95d
DOI: 10.3390/nu12123773 · PMID: 33316914 · PMCID: PMC7764169
782. Vitamin D deficiency as a predictor of poor prognosis in patients with acute respiratory failure due to COVID-19
G. E. Carpagnano, V. Di Lecce, V. N. Quaranta, A. Zito, E. Buonamico, E. Capozza, A. Palumbo, G. Di Gioia, V. N. Valerio, O. Resta Journal of Endocrinological Investigation (2020-08-09) https://doi.org/gg7kqp
DOI: 10.1007/s40618-020-01370-x · PMID: 32772324 · PMCID: PMC7415009
783. Vitamin D Deficiency and Outcome of COVID-19 Patients
Aleksandar Radujkovic, Theresa Hippchen, Shilpa Tiwari-Heckler, Saida Dreher, Monica Boxberger, Uta Merle Nutrients (2020-09-10) https://doi.org/ghgfmp
DOI: 10.3390/nu12092757 · PMID: 32927735 · PMCID: PMC7551780
784. Impact of Vitamin D Deficiency on COVID-19—A Prospective Analysis from the CovILD Registry
Alex Pizzini, Magdalena Aichner, Sabina Sahanic, Anna Böhm, Alexander Egger, Gregor Hoermann, Katharina Kurz, Gerlig Widmann, Rosa Bellmann-Weiler, Günter Weiss, … Judith Löffler-Ragg Nutrients (2020-09-11) https://doi.org/ghr95b
DOI: 10.3390/nu12092775 · PMID: 32932831 · PMCID: PMC7551662
785. Does Serum Vitamin D Level Affect COVID-19 Infection and Its Severity?-A Case-Control Study
Kun Ye, Fen Tang, Xin Liao, Benjamin A. Shaw, Meiqiu Deng, Guangyi Huang, Zhiqiang Qin, Xiaomei Peng, Hewei Xiao, Chunxia Chen, … Jianrong Yang Journal of the American College of Nutrition (2020-10-13) https://doi.org/ghr935
DOI: 10.1080/07315724.2020.1826005 · PMID: 33048028
786. Lower levels of vitamin D are associated with SARS-CoV-2 infection and mortality in the Indian population: An observational study
Sunali Padhi, Subham Suvankar, Venketesh K. Panda, Abhijit Pati, Aditya K. Panda International Immunopharmacology (2020-11) https://doi.org/ghr93w
DOI: 10.1016/j.intimp.2020.107001 · PMID: 33182040 · PMCID: PMC7489890
787. Vitamin D Deficiency Is Associated with COVID-19 Incidence and Disease Severity in Chinese People
Xia Luo, Qing Liao, Ying Shen, Huijun Li, Liming Cheng The Journal of Nutrition (2021-01) https://doi.org/ghr939
DOI: 10.1093/jn/nxaa332 · PMID: 33188401
788. Vitamin D concentrations and COVID-19 infection in UK Biobank
Claire E. Hastie, Daniel F. Mackay, Frederick Ho, Carlos A. Celis-Morales, Srinivasa Vittal Katikireddi, Claire L. Niedzwiedz, Bhautesh D. Jani, Paul Welsh, Frances S. Mair, Stuart R. Gray, … Jill P. Pell Diabetes & Metabolic Syndrome: Clinical Research & Reviews (2020-07) https://doi.org/ggvv72
DOI: 10.1016/j.dsx.2020.04.050 · PMID: 32413819 · PMCID: PMC7204679
790. Low serum 25‐hydroxyvitamin D (25[OH]D) levels in patients hospitalized with COVID‐19 are associated with greater disease severity
Grigorios Panagiotou, Su Ann Tee, Yasir Ihsan, Waseem Athar, Gabriella Marchitelli, Donna Kelly, Christopher S. Boot, Nadia Stock, James Macfarlane, Adrian R. Martineau, … Richard Quinton Clinical Endocrinology (2020-08-06) https://doi.org/gg5gbj
DOI: 10.1111/cen.14276 · PMID: 32621392 · PMCID: PMC7361912
791. Letter in response to the article: Vitamin D concentrations and COVID-19 infection in UK biobank (Hastie et al.)
W. B. Grant, S. L. McDonnell Diabetes & Metabolic Syndrome: Clinical Research & Reviews (2020-09) https://doi.org/ghc7p4
DOI: 10.1016/j.dsx.2020.05.046 · PMID: 32563941 · PMCID: PMC7293469
792. Vitamin D deficiency in African Americans is associated with a high risk of severe disease and mortality by SARS-CoV-2
Virna Margarita Martín Giménez, Felipe Inserra, León Ferder, Joxel García, Walter Manucha Journal of Human Hypertension (2020-08-13) https://doi.org/ghr933
DOI: 10.1038/s41371-020-00398-z · PMID: 32792611 · PMCID: PMC7425793
793. Evidence for possible association of vitamin D status with cytokine storm and unregulated inflammation in COVID-19 patients
Ali Daneshkhah, Vasundhara Agrawal, Adam Eshein, Hariharan Subramanian, Hemant Kumar Roy, Vadim Backman Aging Clinical and Experimental Research (2020-09-02) https://doi.org/ghr93q
DOI: 10.1007/s40520-020-01677-y · PMID: 32876941 · PMCID: PMC7465887
794. Short term, high-dose vitamin D supplementation for COVID-19 disease: a randomised, placebo-controlled, study (SHADE study)
Ashu Rastogi, Anil Bhansali, Niranjan Khare, Vikas Suri, Narayana Yaddanapudi, Naresh Sachdeva, GD Puri, Pankaj Malhotra Postgraduate Medical Journal (2020-11-12) https://doi.org/ghnhpq
DOI: 10.1136/postgradmedj-2020-139065 · PMID: 33184146
795. “Effect of calcifediol treatment and best available therapy versus best available therapy on intensive care unit admission and mortality among patients hospitalized for COVID-19: A pilot randomized clinical study”
Marta Entrenas Castillo, Luis Manuel Entrenas Costa, José Manuel Vaquero Barrios, Juan Francisco Alcalá Díaz, José López Miranda, Roger Bouillon, José Manuel Quesada Gomez The Journal of Steroid Biochemistry and Molecular Biology (2020-10) https://doi.org/ghd79r
DOI: 10.1016/j.jsbmb.2020.105751 · PMID: 32871238 · PMCID: PMC7456194
797. Mathematical analysis of Córdoba calcifediol trial suggests strong role for Vitamin D in reducing ICU admissions of hospitalized COVID-19 patients
Irwin Jungreis, Manolis Kellis Cold Spring Harbor Laboratory (2020-12-21) https://doi.org/ghr94h
DOI: 10.1101/2020.11.08.20222638
798. High-Dose Cholecalciferol Booster Therapy is Associated with a Reduced Risk of Mortality in Patients with COVID-19: A Cross-Sectional Multi-Centre Observational Study
Stephanie F. Ling, Eleanor Broad, Rebecca Murphy, Joseph M. Pappachan, Satveer Pardesi-Newton, Marie-France Kong, Edward B. Jude Nutrients (2020-12-11) https://doi.org/ghr95f
DOI: 10.3390/nu12123799 · PMID: 33322317 · PMCID: PMC7763301
799. Effect of Vitamin D 3 Supplementation vs Placebo on Hospital Length of Stay in Patients with Severe COVID-19: A Multicenter, Double-blind, Randomized Controlled Trial
Igor H. Murai, Alan L. Fernandes, Lucas P. Sales, Ana J. Pinto, Karla F. Goessler, Camila S. C. Duran, Carla B. R. Silva, André S. Franco, Marina B. Macedo, Henrique H. H. Dalmolin, … Rosa M. R. Pereira Cold Spring Harbor Laboratory (2020-11-17) https://doi.org/ghr94j
DOI: 10.1101/2020.11.16.20232397
800. Effect of Vitamin D Administration on Prevention and Treatment of Mild Forms of Suspected Covid-19
Manuel Castillo Garzón clinicaltrials.gov (2020-04-03) https://clinicaltrials.gov/ct2/show/NCT04334005
803. COvid-19 and Vitamin D Supplementation: a Multicenter Randomized Controlled Trial of High Dose Versus Standard Dose Vitamin D3 in High-risk COVID-19 Patients (CoVitTrial) - Full Text View - ClinicalTrials.govhttps://clinicaltrials.gov/ct2/show/NCT04344041
804. The LEAD COVID-19 Trial: Low-risk, Early Aspirin and Vitamin D to Reduce COVID-19 Hospitalizations
Louisiana State University Health Sciences Center in New Orleans clinicaltrials.gov (2020-04-24) https://clinicaltrials.gov/ct2/show/NCT04363840
805. Randomized Double-Blind Placebo-Controlled Proof-of-Concept Trial of a Plant Polyphenol for the Outpatient Treatment of Mild Coronavirus Disease (COVID-19)
Marvin McCreary MD clinicaltrials.gov (2020-09-22) https://clinicaltrials.gov/ct2/show/NCT04400890
806. Current vitamin D status in European and Middle East countries and strategies to prevent vitamin D deficiency: a position statement of the European Calcified Tissue Society
Paul Lips, Kevin D Cashman, Christel Lamberg-Allardt, Heike Annette Bischoff-Ferrari, Barbara Obermayer-Pietsch, Maria Luisa Bianchi, Jan Stepan, Ghada El-Hajj Fuleihan, Roger Bouillon European Journal of Endocrinology (2019-04) https://doi.org/ggr42p
DOI: 10.1530/eje-18-0736 · PMID: 30721133
808. Covid-19: NHS bosses told to assess risk to ethnic minority staff who may be at greater risk
Gareth Iacobucci BMJ (2020-05-04) https://doi.org/ggv2zq
DOI: 10.1136/bmj.m1820 · PMID: 32366503
809. Covid-19: Public health agencies review whether vitamin D supplements could reduce risk
Ingrid Torjesen BMJ (2020-06-19) https://doi.org/ghr94p
DOI: 10.1136/bmj.m2475 · PMID: 32561509
812. Prevalence of Vitamin D Deficiency and Associated Risk Factors in the US Population (2011-2012)
Naveen R Parva, Satish Tadepalli, Pratiksha Singh, Andrew Qian, Rajat Joshi, Hyndavi Kandala, Vinod K Nookala, Pramil Cheriyath Cureus (2018-06-05) https://doi.org/gg7kqq
DOI: 10.7759/cureus.2741 · PMID: 30087817 · PMCID: PMC6075634
816. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic
Colin Hill, Francisco Guarner, Gregor Reid, Glenn R. Gibson, Daniel J. Merenstein, Bruno Pot, Lorenzo Morelli, Roberto Berni Canani, Harry J. Flint, Seppo Salminen, … Mary Ellen Sanders Nature Reviews Gastroenterology & Hepatology (2014-06-10) https://doi.org/f6ndv7
DOI: 10.1038/nrgastro.2014.66 · PMID: 24912386
817. The Effect of Probiotics on Prevention of Common Cold: A Meta-Analysis of Randomized Controlled Trial Studies
En-Jin Kang, Soo Young Kim, In-Hong Hwang, Yun-Jeong Ji Korean Journal of Family Medicine (2013) https://doi.org/gg3knf
DOI: 10.4082/kjfm.2013.34.1.2 · PMID: 23372900 · PMCID: PMC3560336
818. Probiotics and Paraprobiotics in Viral Infection: Clinical Application and Effects on the Innate and Acquired Immune Systems
Osamu Kanauchi, Akira Andoh, Sazaly AbuBakar, Naoki Yamamoto Current Pharmaceutical Design (2018-05-10) https://doi.org/gdjnpk
DOI: 10.2174/1381612824666180116163411 · PMID: 29345577 · PMCID: PMC6006794
819. Using Probiotics to Flatten the Curve of Coronavirus Disease COVID-2019 Pandemic
David Baud, Varvara Dimopoulou Agri, Glenn R. Gibson, Gregor Reid, Eric Giannoni Frontiers in Public Health (2020-05-08) https://doi.org/gg3knd
DOI: 10.3389/fpubh.2020.00186 · PMID: 32574290 · PMCID: PMC7227397
820. Next-generation probiotics: the spectrum from probiotics to live biotherapeutics
Paul W. O’Toole, Julian R. Marchesi, Colin Hill Nature Microbiology (2017-04-25) https://doi.org/ggzggv
DOI: 10.1038/nmicrobiol.2017.57 · PMID: 28440276
823. Probiotic Mechanisms of Action
Miriam Bermudez-Brito, Julio Plaza-Díaz, Sergio Muñoz-Quezada, Carolina Gómez-Llorente, Angel Gil Annals of Nutrition and Metabolism (2012) https://doi.org/gg3knb
DOI: 10.1159/000342079 · PMID: 23037511
824. A novel eukaryotic cell culture model to study antiviral activity of potential probiotic bacteria
T BOTIC, T KLINGBERG, H WEINGARTL, A CENCIC International Journal of Food Microbiology (2007-04-30) https://doi.org/fks5cz
DOI: 10.1016/j.ijfoodmicro.2006.10.044 · PMID: 17261339
825. Oral administration of Lactobacillus brevis KB290 to mice alleviates clinical symptoms following influenza virus infection
N. Waki, N. Yajima, H. Suganuma, B. M. Buddle, D. Luo, A. Heiser, T. Zheng Letters in Applied Microbiology (2014-01) https://doi.org/f5j37w
DOI: 10.1111/lam.12160 · PMID: 24329975
826. Antiviral activity of Lactobacillus brevis towards herpes simplex virus type 2: Role of cell wall associated components
Paola Mastromarino, Fatima Cacciotti, Alessandra Masci, Luciana Mosca Anaerobe (2011-12) https://doi.org/bcpvm5
DOI: 10.1016/j.anaerobe.2011.04.022 · PMID: 21621625
827. Critical Adverse Impact of IL-6 in Acute Pneumovirus Infection
Caroline M. Percopo, Michelle Ma, Todd A. Brenner, Julia O. Krumholz, Timothy J. Break, Karen Laky, Helene F. Rosenberg The Journal of Immunology (2019-02-01) https://doi.org/ghr95h
DOI: 10.4049/jimmunol.1800927 · PMID: 30578308 · PMCID: PMC6365009
828. Antiviral Activity of Exopolysaccharides Produced by Lactic Acid Bacteria of the Genera Pediococcus, Leuconostoc and Lactobacillus against Human Adenovirus Type 5
Biliavska, Pankivska, Povnitsa, Zagorodnya Medicina (2019-08-22) https://doi.org/ghr948
DOI: 10.3390/medicina55090519 · PMID: 31443536 · PMCID: PMC6780409
830. Effect of probiotic on innate inflammatory response and viral shedding in experimental rhinovirus infection – a randomised controlled trial
R. B. Turner, J. A. Woodfolk, L. Borish, J. W. Steinke, J. T. Patrie, L. M. Muehling, S. Lahtinen, M. J. Lehtinen Beneficial Microbes (2017-04-26) https://doi.org/f955fh
DOI: 10.3920/bm2016.0160 · PMID: 28343401 · PMCID: PMC5797652
831. Immunobiotic lactobacilli reduce viral-associated pulmonary damage through the modulation of inflammation–coagulation interactions
Hortensia Zelaya, Kohichiro Tsukida, Eriko Chiba, Gabriela Marranzino, Susana Alvarez, Haruki Kitazawa, Graciela Agüero, Julio Villena International Immunopharmacology (2014-03) https://doi.org/f5wd93
DOI: 10.1016/j.intimp.2013.12.020 · PMID: 24394565
832. Nasal priming with immunobiotic lactobacilli improves the adaptive immune response against influenza virus
Fernanda Raya Tonetti, Md. Aminul Islam, Maria Guadalupe Vizoso-Pinto, Hideki Takahashi, Haruki Kitazawa, Julio Villena International Immunopharmacology (2020-01) https://doi.org/ghr93v
DOI: 10.1016/j.intimp.2019.106115 · PMID: 31841753
833. The potential application of probiotics and prebiotics for the prevention and treatment of COVID-19
Amin N. Olaimat, Iman Aolymat, Murad Al-Holy, Mutamed Ayyash, Mahmoud Abu Ghoush, Anas A. Al-Nabulsi, Tareq Osaili, Vasso Apostolopoulos, Shao-Quan Liu, Nagendra P. Shah npj Science of Food (2020-10-05) https://doi.org/ghggq4
DOI: 10.1038/s41538-020-00078-9 · PMID: 33083549 · PMCID: PMC7536434
835. The role of the lung microbiota and the gut-lung axis in respiratory infectious diseases
Alexia Dumas, Lucie Bernard, Yannick Poquet, Geanncarlo Lugo-Villarino, Olivier Neyrolles Cellular Microbiology (2018-12) https://doi.org/gfjds9
DOI: 10.1111/cmi.12966 · PMID: 30329198
841. Probiotics for the prevention of respiratory tract infections: a systematic review
Evridiki K. Vouloumanou, Gregory C. Makris, Drosos E. Karageorgopoulos, Matthew E. Falagas International Journal of Antimicrobial Agents (2009-09) https://doi.org/dn8kw8
DOI: 10.1016/j.ijantimicag.2008.11.005 · PMID: 19179052
842. Effectiveness of probiotics on the duration of illness in healthy children and adults who develop common acute respiratory infectious conditions: a systematic review and meta-analysis
Sarah King, Julie Glanville, Mary Ellen Sanders, Anita Fitzgerald, Danielle Varley British Journal of Nutrition (2014-04-29) https://doi.org/f57hq5
DOI: 10.1017/s0007114514000075 · PMID: 24780623 · PMCID: PMC4054664
843. Effect of probiotics on the incidence of ventilator-associated pneumonia in critically ill patients: a randomized controlled multicenter trial
Juan Zeng, Chun-Ting Wang, Fu-Shen Zhang, Feng Qi, Shi-Fu Wang, Shuang Ma, Tie-Jun Wu, Hui Tian, Zhao-Tao Tian, Shu-Liu Zhang, … Yu-Ping Wang Intensive Care Medicine (2016-04-04) https://doi.org/f8jnrt
DOI: 10.1007/s00134-016-4303-x · PMID: 27043237
844. Probiotic Prophylaxis of Ventilator-associated Pneumonia
Lee E. Morrow, Marin H. Kollef, Thomas B. Casale American Journal of Respiratory and Critical Care Medicine (2010-10-15) https://doi.org/d5hh4t
DOI: 10.1164/rccm.200912-1853oc · PMID: 20522788 · PMCID: PMC2970846
845. Synbiotics modulate gut microbiota and reduce enteritis and ventilator-associated pneumonia in patients with sepsis: a randomized controlled trial
Kentaro Shimizu, Tomoki Yamada, Hiroshi Ogura, Tomoyoshi Mohri, Takeyuki Kiguchi, Satoshi Fujimi, Takashi Asahara, Tomomi Yamada, Masahiro Ojima, Mitsunori Ikeda, Takeshi Shimazu Critical Care (2018-09-27) https://doi.org/gfdggj
DOI: 10.1186/s13054-018-2167-x · PMID: 30261905 · PMCID: PMC6161427
846. Probiotics for the Prevention of Ventilator-Associated Pneumonia: A Meta-Analysis of Randomized Controlled Trials
Minmin Su, Ying Jia, Yan Li, Dianyou Zhou, Jinsheng Jia Respiratory Care (2020-05) https://doi.org/gg3kng
DOI: 10.4187/respcare.07097 · PMID: 32127415
848. The challenge of ventilator-associated pneumonia diagnosis in COVID-19 patients
Bruno François, Pierre-François Laterre, Charles-Edouard Luyt, Jean Chastre Critical Care (2020-06-05) https://doi.org/gg3knc
DOI: 10.1186/s13054-020-03013-2 · PMID: 32503590 · PMCID: PMC7273812
849. Prophylactic use of probiotics for gastrointestinal disorders in children
Celine Perceval, Hania Szajewska, Flavia Indrio, Zvi Weizman, Yvan Vandenplas The Lancet Child & Adolescent Health (2019-09) https://doi.org/d2qp
DOI: 10.1016/s2352-4642(19)30182-8
851. The digestive system is a potential route of 2019-nCov infection: a bioinformatics analysis based on single-cell transcriptomes
Hao Zhang, Zijian Kang, Haiyi Gong, Da Xu, Jing Wang, Zifu Li, Xingang Cui, Jianru Xiao, Tong Meng, Wang Zhou, … Huji Xu Cold Spring Harbor Laboratory (2020-01-31) https://doi.org/ggjvx2
DOI: 10.1101/2020.01.30.927806
852. Cryo-electron microscopy structures of the SARS-CoV spike glycoprotein reveal a prerequisite conformational state for receptor binding
Miao Gui, Wenfei Song, Haixia Zhou, Jingwei Xu, Silian Chen, Ye Xiang, Xinquan Wang Cell Research (2016-12-23) https://doi.org/f9m247
DOI: 10.1038/cr.2016.152 · PMID: 28008928 · PMCID: PMC5223232
854. Enteric involvement of coronaviruses: is faecal–oral transmission of SARS-CoV-2 possible?
Charleen Yeo, Sanghvi Kaushal, Danson Yeo The Lancet Gastroenterology & Hepatology (2020-04) https://doi.org/ggpx7s
DOI: 10.1016/s2468-1253(20)30048-0 · PMID: 32087098 · PMCID: PMC7130008
855. Characteristics of pediatric SARS-CoV-2 infection and potential evidence for persistent fecal viral shedding
Yi Xu, Xufang Li, Bing Zhu, Huiying Liang, Chunxiao Fang, Yu Gong, Qiaozhi Guo, Xin Sun, Danyang Zhao, Jun Shen, … Sitang Gong Nature Medicine (2020-03-13) https://doi.org/ggpwx5
DOI: 10.1038/s41591-020-0817-4 · PMID: 32284613 · PMCID: PMC7095102
856. Modulation of rotavirus severe gastroenteritis by the combination of probiotics and prebiotics
Guadalupe Gonzalez-Ochoa, Lilian K. Flores-Mendoza, Ramona Icedo-Garcia, Ricardo Gomez-Flores, Patricia Tamez-Guerra Archives of Microbiology (2017-06-20) https://doi.org/gbsb4d
DOI: 10.1007/s00203-017-1400-3 · PMID: 28634691 · PMCID: PMC5548957
857. Multicenter Trial of a Combination Probiotic for Children with Gastroenteritis
Stephen B. Freedman, Sarah Williamson-Urquhart, Ken J. Farion, Serge Gouin, Andrew R. Willan, Naveen Poonai, Katrina Hurley, Philip M. Sherman, Yaron Finkelstein, Bonita E. Lee, … Suzanne Schuh New England Journal of Medicine (2018-11-22) https://doi.org/gfkbsf
DOI: 10.1056/nejmoa1802597 · PMID: 30462939
858. Synbiotic Therapy of Gastrointestinal Symptoms During Covid-19 Infection: A Randomized, Double-blind, Placebo Controlled, Telemedicine Study (SynCov Study)
Medical University of Graz clinicaltrials.gov (2021-01-14) https://clinicaltrials.gov/ct2/show/NCT04420676
859. Multicentric Study to Assess the Effect of Consumption of Lactobacillus Coryniformis K8 on Healthcare Personnel Exposed to COVID-19
Biosearch S.A. clinicaltrials.gov (2020-04-28) https://clinicaltrials.gov/ct2/show/NCT04366180
860. The Intestinal Microbiota as a Therapeutic Target in Hospitalized Patients With COVID-19 Infection
Bioithas SL clinicaltrials.gov (2021-01-26) https://clinicaltrials.gov/ct2/show/NCT04390477
862. Health benefits and health claims of probiotics: bridging science and marketing
Ger T. Rijkers, Willem M. de Vos, Robert-Jan Brummer, Lorenzo Morelli, Gerard Corthier, Philippe Marteau British Journal of Nutrition (2011-08-24) https://doi.org/cb78rx
DOI: 10.1017/s000711451100287x · PMID: 21861940
868. Obesogenic diet in aging mice disrupts gut microbe composition and alters neutrophi:lymphocyte ratio, leading to inflamed milieu in acute heart failure
Vasundhara Kain, William Van Der Pol, Nithya Mariappan, Aftab Ahmad, Peter Eipers, Deanna L. Gibson, Cecile Gladine, Claire Vigor, Thierry Durand, Casey Morrow, Ganesh V. Halade The FASEB Journal (2019-02-15) https://doi.org/ghwfq8
DOI: 10.1096/fj.201802477r · PMID: 30768364 · PMCID: PMC6463911
871. Role of Melatonin on Virus-Induced Neuropathogenesis—A Concomitant Therapeutic Strategy to Understand SARS-CoV-2 Infection
Prapimpun Wongchitrat, Mayuri Shukla, Ramaswamy Sharma, Piyarat Govitrapong, Russel J. Reiter Antioxidants (2021-01-02) https://doi.org/ghr946
DOI: 10.3390/antiox10010047 · PMID: 33401749 · PMCID: PMC7823793
872. Nutraceutical Strategies for Suppressing NLRP3 Inflammasome Activation: Pertinence to the Management of COVID-19 and Beyond
Mark F. McCarty, Simon Bernard Iloki Assanga, Lidianys Lewis Luján, James H. O’Keefe, James J. DiNicolantonio Nutrients (2020-12-25) https://doi.org/ghr95g
DOI: 10.3390/nu13010047 · PMID: 33375692 · PMCID: PMC7823562
874. Dietary supplements during the COVID-19 pandemic: insights from 1.4M users of the COVID Symptom Study app - a longitudinal app-based community survey
Panayiotis Louca, Benjamin Murray, Kerstin Klaser, Mark S Graham, Mohsen Mazidi, Emily R Leeming, Ellen Thompson, Ruth Bowyer, David A Drew, Long H Nguyen, … Cristina Menni Cold Spring Harbor Laboratory (2020-11-30) https://doi.org/ghr94k
DOI: 10.1101/2020.11.27.20239087
875. ESPEN expert statements and practical guidance for nutritional management of individuals with SARS-CoV-2 infection
Rocco Barazzoni, Stephan C. Bischoff, Joao Breda, Kremlin Wickramasinghe, Zeljko Krznaric, Dorit Nitzan, Matthias Pirlich, Pierre Singer Clinical Nutrition (2020-06) https://doi.org/ggtzjq
DOI: 10.1016/j.clnu.2020.03.022 · PMID: 32305181 · PMCID: PMC7138149
876. Nutritional status assessment in patients with Covid-19 after discharge from the intensive care unit
Nassim Essabah Haraj, Siham El Aziz, Asma Chadli, Asma Dafir, Amal Mjabber, Ouissal Aissaoui, Lhoucine Barrou, Chafik El Kettani El Hamidi, Afak Nsiri, Rachid AL Harrar, … Moulay Hicham Afif Clinical Nutrition ESPEN (2021-02) https://doi.org/ghjhdq
DOI: 10.1016/j.clnesp.2020.09.214 · PMID: 33487301 · PMCID: PMC7552965
878. Evaluation of Nutrition Risk and Its Association With Mortality Risk in Severely and Critically Ill COVID‐19 Patients
Xiaobo Zhao, Yan Li, Yanyan Ge, Yuxin Shi, Ping Lv, Jianchu Zhang, Gui Fu, Yanfen Zhou, Ke Jiang, Nengxing Lin, … Xin Li Journal of Parenteral and Enteral Nutrition (2020-07-20) https://doi.org/ghr93n
DOI: 10.1002/jpen.1953 · PMID: 32613660 · PMCID: PMC7361906
879. Multisystem inflammatory syndrome in children: A systematic review
Mubbasheer Ahmed, Shailesh Advani, Axel Moreira, Sarah Zoretic, John Martinez, Kevin Chorath, Sebastian Acosta, Rija Naqvi, Finn Burmeister-Morton, Fiona Burmeister, … Alvaro Moreira EClinicalMedicine (2020-09) https://doi.org/ghsv27
DOI: 10.1016/j.eclinm.2020.100527 · PMID: 32923992 · PMCID: PMC7473262
880. Nutritional management of COVID-19 patients in a rehabilitation unit
Luigia Brugliera, Alfio Spina, Paola Castellazzi, Paolo Cimino, Pietro Arcuri, Alessandra Negro, Elise Houdayer, Federica Alemanno, Alessandra Giordani, Pietro Mortini, Sandro Iannaccone European Journal of Clinical Nutrition (2020-05-20) https://doi.org/gg29hf
DOI: 10.1038/s41430-020-0664-x · PMID: 32433599 · PMCID: PMC7237874
881. The frontier between nutrition and pharma: The international regulatory framework of functional foods, food supplements and nutraceuticals
Laura Domínguez Díaz, Virginia Fernández-Ruiz, Montaña Cámara Critical Reviews in Food Science and Nutrition (2019-03-29) https://doi.org/ggqs3w
DOI: 10.1080/10408398.2019.1592107 · PMID: 30924346
884. Potential roles of social distancing in mitigating the spread of coronavirus disease 2019 (COVID-19) in South Korea
Sang Woo Park, Kaiyuan Sun, Cécile Viboud, Bryan T. Grenfell, Jonathan Dushoff Cold Spring Harbor Laboratory (2020-03-30) https://doi.org/gg3mhg
DOI: 10.1101/2020.03.27.20045815 · PMID: 32511429 · PMCID: PMC7217070
885. Evaluating the Effectiveness of Social Distancing Interventions to Delay or Flatten the Epidemic Curve of Coronavirus Disease
Laura Matrajt, Tiffany Leung Emerging Infectious Diseases (2020-08) https://doi.org/ggtx3k
DOI: 10.3201/eid2608.201093 · PMID: 32343222 · PMCID: PMC7392458
887. Developing Covid-19 Vaccines at Pandemic Speed
Nicole Lurie, Melanie Saville, Richard Hatchett, Jane Halton New England Journal of Medicine (2020-05-21) https://doi.org/ggq8bc
DOI: 10.1056/nejmp2005630 · PMID: 32227757
894. An mRNA Vaccine against SARS-CoV-2 — Preliminary Report
Lisa A. Jackson, Evan J. Anderson, Nadine G. Rouphael, Paul C. Roberts, Mamodikoe Makhene, Rhea N. Coler, Michele P. McCullough, James D. Chappell, Mark R. Denison, Laura J. Stevens, … John H. Beigel New England Journal of Medicine (2020-11-12) https://doi.org/d3tt
DOI: 10.1056/nejmoa2022483 · PMID: 32663912 · PMCID: PMC7377258
898. Tolerability of intramuscular and intradermal delivery by CELLECTRA ® adaptive constant current electroporation device in healthy volunteers
Malissa C Diehl, Jessica C Lee, Stephen E Daniels, Pablo Tebas, Amir S Khan, Mary Giffear, Niranjan Y Sardesai, Mark L Bagarazzi Human Vaccines & Immunotherapeutics (2014-10-27) https://doi.org/ggrj7h
DOI: 10.4161/hv.24702 · PMID: 24051434 · PMCID: PMC3906411
899. Induction of virus-specific cytotoxic T lymphocytesin vivo by liposome-entrapped mRNA
Frédéric Martinon, Sivadasan Krishnan, Gerlinde Lenzen, Rémy Magné, Elisabeth Gomard, Jean-Gérard Guillet, Jean-Paul Lévy, Pierre Meulien European Journal of Immunology (1993-07) https://doi.org/b6jb3z
DOI: 10.1002/eji.1830230749 · PMID: 8325342
901. mRNA vaccine delivery using lipid nanoparticles
Andreas M Reichmuth, Matthias A Oberli, Ana Jaklenec, Robert Langer, Daniel Blankschtein Therapeutic Delivery (2016-05) https://doi.org/f8xfzc
DOI: 10.4155/tde-2016-0006 · PMID: 27075952 · PMCID: PMC5439223
902. Mechanism of action of mRNA-based vaccines
Carlo Iavarone, Derek T. O’hagan, Dong Yu, Nicolas F. Delahaye, Jeffrey B. Ulmer Expert Review of Vaccines (2017-07-28) https://doi.org/ggsnm6
DOI: 10.1080/14760584.2017.1355245 · PMID: 28701102
904. Study in Healthy Adults to Evaluate Gene Activation After Vaccination With GlaxoSmithKline (GSK) Biologicals’ Candidate Tuberculosis (TB) Vaccine GSK 692342 - Full Text View - ClinicalTrials.govhttps://clinicaltrials.gov/ct2/show/NCT01669096
905. Nucleoside-modified mRNA immunization elicits influenza virus hemagglutinin stalk-specific antibodies
Norbert Pardi, Kaela Parkhouse, Ericka Kirkpatrick, Meagan McMahon, Seth J. Zost, Barbara L. Mui, Ying K. Tam, Katalin Karikó, Christopher J. Barbosa, Thomas D. Madden, … Drew Weissman Nature Communications (2018-08-22) https://doi.org/gd49qt
DOI: 10.1038/s41467-018-05482-0 · PMID: 30135514 · PMCID: PMC6105651
906. Cell specific delivery of modified mRNA expressing therapeutic proteins to leukocytes
Nuphar Veiga, Meir Goldsmith, Yasmin Granot, Daniel Rosenblum, Niels Dammes, Ranit Kedmi, Srinivas Ramishetti, Dan Peer Nature Communications (2018-10-29) https://doi.org/gfmcrt
DOI: 10.1038/s41467-018-06936-1 · PMID: 30374059 · PMCID: PMC6206083
909. mRNA vaccines — a new era in vaccinology
Norbert Pardi, Michael J. Hogan, Frederick W. Porter, Drew Weissman Nature Reviews Drug Discovery (2018-01-12) https://doi.org/gcsmgr
DOI: 10.1038/nrd.2017.243 · PMID: 29326426 · PMCID: PMC5906799
910. A Snapshot of the Global Race for Vaccines Targeting SARS-CoV-2 and the COVID-19 Pandemic
Colin D. Funk, Craig Laferrière, Ali Ardakani Frontiers in Pharmacology (2020-06-19) https://doi.org/gg4hxd
DOI: 10.3389/fphar.2020.00937 · PMID: 32636754 · PMCID: PMC7317023
911. Phase I, Open-Label, Dose-Ranging Study of the Safety and Immunogenicity of 2019-nCoV Vaccine (mRNA-1273) in Healthy Adults
National Institute of Allergy and Infectious Diseases (NIAID) clinicaltrials.gov (2020-12-17) https://clinicaltrials.gov/ct2/show/NCT04283461
914. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine
Lindsey R. Baden, Hana M. El Sahly, Brandon Essink, Karen Kotloff, Sharon Frey, Rick Novak, David Diemert, Stephen A. Spector, Nadine Rouphael, C. Buddy Creech, … Tal Zaks New England Journal of Medicine (2020-12-30) https://doi.org/ghrg8m
DOI: 10.1056/nejmoa2035389 · PMID: 33378609 · PMCID: PMC7787219
915. Vaccines and Related Biological Products Advisory Committee December 17, 2020 Meeting Briefing Document - FDA
FDA/CBER
(2020-12-15) https://www.fda.gov/media/144434/download
922. Synthetic Chemically Modified mRNA (modRNA): Toward a New Technology Platform for Cardiovascular Biology and Medicine
K. R. Chien, L. Zangi, K. O. Lui Cold Spring Harbor Perspectives in Medicine (2014-10-09) https://doi.org/f3pvsr
DOI: 10.1101/cshperspect.a014035 · PMID: 25301935 · PMCID: PMC4292072
924. Phase I/II study of COVID-19 RNA vaccine BNT162b1 in adults
Mark J. Mulligan, Kirsten E. Lyke, Nicholas Kitchin, Judith Absalon, Alejandra Gurtman, Stephen Lockhart, Kathleen Neuzil, Vanessa Raabe, Ruth Bailey, Kena A. Swanson, … Kathrin U. Jansen Nature (2020-08-12) https://doi.org/gg7ww9
DOI: 10.1038/s41586-020-2639-4 · PMID: 32785213
925. COVID-19 vaccine BNT162b1 elicits human antibody and TH1 T cell responses
Ugur Sahin, Alexander Muik, Evelyna Derhovanessian, Isabel Vogler, Lena M. Kranz, Mathias Vormehr, Alina Baum, Kristen Pascal, Jasmin Quandt, Daniel Maurus, … Özlem Türeci Nature (2020-09-30) https://doi.org/ghfmb2
DOI: 10.1038/s41586-020-2814-7 · PMID: 32998157
932. Development and Applications of Viral Vectored Vaccines to Combat Zoonotic and Emerging Public Health Threats
Sophia M. Vrba, Natalie M. Kirk, Morgan E. Brisse, Yuying Liang, Hinh Ly Vaccines (2020-11-13) https://doi.org/gh23ww
DOI: 10.3390/vaccines8040680 · PMID: 33202961 · PMCID: PMC7712223
933. Viral vectors as vaccine platforms: deployment in sight
Christine S Rollier, Arturo Reyes-Sandoval, Matthew G Cottingham, Katie Ewer, Adrian VS Hill Current Opinion in Immunology (2011-06) https://doi.org/d8p72q
DOI: 10.1016/j.coi.2011.03.006 · PMID: 21514130
939. International seroepidemiology of adenovirus serotypes 5, 26, 35, and 48 in pediatric and adult populations
Dan H. Barouch, Sandra V. Kik, Gerrit J. Weverling, Rebecca Dilan, Sharon L. King, Lori F. Maxfield, Sarah Clark, David Ng’ang’a, Kara L. Brandariz, Peter Abbink, … Jaap Goudsmit Vaccine (2011-07) https://doi.org/bmzpdx
DOI: 10.1016/j.vaccine.2011.05.025 · PMID: 21619905 · PMCID: PMC3138857
941. Safety and immunogenicity of an rAd26 and rAd5 vector-based heterologous prime-boost COVID-19 vaccine in two formulations: two open, non-randomised phase 1/2 studies from Russia
Denis Y Logunov, Inna V Dolzhikova, Olga V Zubkova, Amir I Tukhvatulin, Dmitry V Shcheblyakov, Alina S Dzharullaeva, Daria M Grousova, Alina S Erokhova, Anna V Kovyrshina, Andrei G Botikov, … Alexander L Gintsburg The Lancet (2020-09) https://doi.org/gg96hq
DOI: 10.1016/s0140-6736(20)31866-3 · PMID: 32896291 · PMCID: PMC7471804
942. Lead SARS-CoV-2 Candidate Vaccines: Expectations from Phase III Trials and Recommendations Post-Vaccine Approval
Ebenezer Tumban Viruses (2020-12-31) https://doi.org/gh2z7h
DOI: 10.3390/v13010054 · PMID: 33396343 · PMCID: PMC7824305
947. Safety and efficacy of an rAd26 and rAd5 vector-based heterologous prime-boost COVID-19 vaccine: an interim analysis of a randomised controlled phase 3 trial in Russia
Denis Y Logunov, Inna V Dolzhikova, Dmitry V Shcheblyakov, Amir I Tukhvatulin, Olga V Zubkova, Alina S Dzharullaeva, Anna V Kovyrshina, Nadezhda L Lubenets, Daria M Grousova, Alina S Erokhova, … Alexander L Gintsburg The Lancet (2021-02) https://doi.org/ghxj4g
DOI: 10.1016/s0140-6736(21)00234-8 · PMID: 33545094 · PMCID: PMC7852454
951. ACTIVE SURVEILLANCE OF THE SPUTNIK V VACCINE IN HEALTH WORKERS
Vanina Pagotto, Analia Ferloni, María Mercedes Soriano, Morena Díaz, Manuel Braguisnky Golde, María Isabel González, Valeria Asprea, Inés Staneloni, Gustavo Vidal, Martin Silveira, … Silvana Figar medRxiv (2021-02-05) https://www.medrxiv.org/content/10.1101/2021.02.03.21251071v1
DOI: 10.1101/2021.02.03.21251071
954. Study in Adults to Determine the Safety and Immunogenicity of AZD1222, a Non-replicating ChAdOx1 Vector Vaccine, Given in Combination With rAd26-S, Recombinant Adenovirus Type 26 Component of Gam-COVID-Vac Vaccine, for the Prevention of COVID-19. - Full Text View - ClinicalTrials.govhttps://clinicaltrials.gov/ct2/show/NCT04686773
956. Operation Warp Speed: Accelerated COVID-19 Vaccine Development Status and Efforts to Address Manufacturing Challenges
U. S. Government Accountability Office https://www.gao.gov/products/gao-21-319
958. Interim Results of a Phase 1–2a Trial of Ad26.COV2.S Covid-19 Vaccine
Jerald Sadoff, Mathieu Le Gars, Georgi Shukarev, Dirk Heerwegh, Carla Truyers, Anne M. de Groot, Jeroen Stoop, Sarah Tete, Wim Van Damme, Isabel Leroux-Roels, … Hanneke Schuitemaker New England Journal of Medicine (2021-01-13) https://doi.org/fqnt
DOI: 10.1056/nejmoa2034201 · PMID: 33440088 · PMCID: PMC7821985
959. Ad26 vector-based COVID-19 vaccine encoding a prefusion-stabilized SARS-CoV-2 Spike immunogen induces potent humoral and cellular immune responses
Rinke Bos, Lucy Rutten, Joan E. M. van der Lubbe, Mark J. G. Bakkers, Gijs Hardenberg, Frank Wegmann, David Zuijdgeest, Adriaan H. de Wilde, Annemart Koornneef, Annemiek Verwilligen, … Hanneke Schuitemaker npj Vaccines (2020-09-28) https://doi.org/ghjkr8
DOI: 10.1038/s41541-020-00243-x · PMID: 33083026 · PMCID: PMC7522255
961. Single-shot Ad26 vaccine protects against SARS-CoV-2 in rhesus macaques
Noe B. Mercado, Roland Zahn, Frank Wegmann, Carolin Loos, Abishek Chandrashekar, Jingyou Yu, Jinyan Liu, Lauren Peter, Katherine McMahan, Lisa H. Tostanoski, … Dan H. Barouch Nature (2020-07-30) https://doi.org/d5d4
DOI: 10.1038/s41586-020-2607-z · PMID: 32731257 · PMCID: PMC7581548
962. Ad26 vaccine protects against SARS-CoV-2 severe clinical disease in hamsters
Lisa H. Tostanoski, Frank Wegmann, Amanda J. Martinot, Carolin Loos, Katherine McMahan, Noe B. Mercado, Jingyou Yu, Chi N. Chan, Stephen Bondoc, Carly E. Starke, … Dan H. Barouch Nature Medicine (2020-11) https://doi.org/gjhgd2
DOI: 10.1038/s41591-020-1070-6 · PMID: 32884153 · PMCID: PMC7671939
963. Immunogenicity and protective efficacy of one- and two-dose regimens of the Ad26.COV2.S COVID-19 vaccine candidate in adult and aged rhesus macaques
Laura Solforosi, Harmjan Kuipers, Sietske K. Rosendahl Huber, Joan E. M. van der Lubbe, Liesbeth Dekking, Dominika N. Czapska-Casey, Ana Izquierdo Gil, Miranda R. M. Baert, Joke Drijver, Joost Vaneman, … Roland C. Zahn Immunology (2020-11-17) https://doi.org/ghwzk9
DOI: 10.1101/2020.11.17.368258
964. Low-Dose Ad26.COV2.S Protection Against SARS-CoV-2 Challenge in Rhesus Macaques
Xuan He, Abishek Chandrashekar, Roland Zahn, Frank Wegmann, Jingyou Yu, Noe B. Mercado, Katherine McMahan, Amanda J. Martinot, Cesar Piedra-Mora, Sidney Beecy, … Dan H. Barouch Cold Spring Harbor Laboratory (2021-01-27) https://doi.org/gjhgd3
DOI: 10.1101/2021.01.27.428380 · PMID: 33532782 · PMCID: PMC7852276
965. SARS-CoV-2 binding and neutralizing antibody levels after vaccination with Ad26.COV2.S predict durable protection in rhesus macaques
Ramon Roozendaal, Laura Solforosi, Daniel Stieh, Jan Serroyen, Roel Straetemans, Frank Wegmann, Sietske K. Rosendahl Huber, Joan E. M. van der Lubbe, Jenny Hendriks, Mathieu le Gars, … Roland Zahn Immunology (2021-01-30) https://doi.org/gjhgd4
DOI: 10.1101/2021.01.30.428921
966. Safety and immunogenicity of the Ad26.COV2.S COVID-19 vaccine candidate: interim results of a phase 1/2a, double-blind, randomized, placebo-controlled trial
Jerald Sadoff, Mathieu Le Gars, Georgi Shukarev, Dirk Heerwegh, Carla Truyers, Anne Marit de Groot, Jeroen Stoop, Sarah Tete, Wim Van Damme, Isabel Leroux-Roels, … Hanneke Schuitemaker Cold Spring Harbor Laboratory (2020-09-25) https://doi.org/ghjk2q
DOI: 10.1101/2020.09.23.20199604
967. Immunogenicity of the Ad26.COV2.S Vaccine for COVID-19
Kathryn E. Stephenson, Mathieu Le Gars, Jerald Sadoff, Anne Marit de Groot, Dirk Heerwegh, Carla Truyers, Caroline Atyeo, Carolin Loos, Abishek Chandrashekar, Katherine McMahan, … Dan H. Barouch JAMA (2021-03-11) https://doi.org/gjhgdz
DOI: 10.1001/jama.2021.3645 · PMID: 33704352 · PMCID: PMC7953339
968. Correlates of protection against SARS-CoV-2 in rhesus macaques
Katherine McMahan, Jingyou Yu, Noe B. Mercado, Carolin Loos, Lisa H. Tostanoski, Abishek Chandrashekar, Jinyan Liu, Lauren Peter, Caroline Atyeo, Alex Zhu, … Dan H. Barouch Nature (2020-12-04) https://doi.org/fmjk
DOI: 10.1038/s41586-020-03041-6 · PMID: 33276369 · PMCID: PMC7906955
969. A Randomized, Double-blind, Placebo-controlled Phase 3 Study to Assess the Efficacy and Safety of Ad26.COV2.S for the Prevention of SARS-CoV-2-mediated COVID-19 in Adults Aged 18 Years and Older
Janssen Vaccines & Prevention B.V. clinicaltrials.gov (2021-03-24) https://clinicaltrials.gov/ct2/show/NCT04505722
974. Vero cell technology for rapid development of inactivated whole virus vaccines for emerging viral diseases
P. Noel Barrett, Sara J. Terpening, Doris Snow, Ronald R. Cobb, Otfried Kistner Expert Review of Vaccines (2017-07-27) https://doi.org/ggt7vf
DOI: 10.1080/14760584.2017.1357471 · PMID: 28724343
975. Vaccine delivery: a matter of size, geometry, kinetics and molecular patterns
Martin F. Bachmann, Gary T. Jennings Nature Reviews Immunology (2010-10-15) https://doi.org/fg5dx9
DOI: 10.1038/nri2868 · PMID: 20948547
976. Functional analysis of influenza-specific helper T cell clones in vivo. T cells specific for internal viral proteins provide cognate help for B cell responses to hemagglutinin.
PA Scherle, W Gerhard Journal of Experimental Medicine (1986-10-01) https://doi.org/bp47qh
DOI: 10.1084/jem.164.4.1114 · PMID: 2944982 · PMCID: PMC2188433
977. A Review of the Progress and Challenges of Developing a Vaccine for COVID-19
Omna Sharma, Ali A. Sultan, Hong Ding, Chris R. Triggle Frontiers in Immunology (2020-10-14) https://doi.org/gh65wd
DOI: 10.3389/fimmu.2020.585354 · PMID: 33163000 · PMCID: PMC7591699
978. Development of an inactivated vaccine candidate for SARS-CoV-2
Qiang Gao, Linlin Bao, Haiyan Mao, Lin Wang, Kangwei Xu, Minnan Yang, Yajing Li, Ling Zhu, Nan Wang, Zhe Lv, … Chuan Qin Science (2020-07-03) https://doi.org/ggvckc
DOI: 10.1126/science.abc1932 · PMID: 32376603 · PMCID: PMC7202686
979. Safety, tolerability, and immunogenicity of an inactivated SARS-CoV-2 vaccine in healthy adults aged 18–59 years: a randomised, double-blind, placebo-controlled, phase 1/2 clinical trial
Yanjun Zhang, Gang Zeng, Hongxing Pan, Changgui Li, Yaling Hu, Kai Chu, Weixiao Han, Zhen Chen, Rong Tang, Weidong Yin, … Fengcai Zhu The Lancet Infectious Diseases (2021-02) https://doi.org/fpcx
DOI: 10.1016/s1473-3099(20)30843-4 · PMID: 33217362 · PMCID: PMC7832443
980. Safety, tolerability, and immunogenicity of an inactivated SARS-CoV-2 vaccine (CoronaVac) in healthy adults aged 60 years and older: a randomised, double-blind, placebo-controlled, phase 1/2 clinical trial
Zhiwei Wu, Yaling Hu, Miao Xu, Zhen Chen, Wanqi Yang, Zhiwei Jiang, Minjie Li, Hui Jin, Guoliang Cui, Panpan Chen, … Weidong Yin The Lancet Infectious Diseases (2021-02) https://doi.org/fx8z
DOI: 10.1016/s1473-3099(20)30987-7 · PMID: 33548194 · PMCID: PMC7906628
981. Safety and immunogenicity of a novel human Enterovirus 71 (EV71) vaccine: A randomized, placebo-controlled, double-blind, Phase I clinical trial
Yan-Ping Li, Zheng-Lun Liang, Qiang Gao, Li-Rong Huang, Qun-Ying Mao, Shu-Qun Wen, Yan Liu, Wei-Dong Yin, Rong-Cheng Li, Jun-Zhi Wang Vaccine (2012-05) https://doi.org/gh7tjn
DOI: 10.1016/j.vaccine.2012.03.010 · PMID: 22426327
982. Double-Blind, Randomized, Placebo-Controlled Phase III Clinical Trial to Evaluate the Efficacy and Safety of treating Healthcare Professionals with the Adsorbed COVID-19 (Inactivated) Vaccine Manufactured by Sinovac – PROFISCOV: A structured summary of a study protocol for a randomised controlled trial
Ricardo Palacios, Elizabeth González Patiño, Roberta de Oliveira Piorelli, Monica Tilli Reis Pessoa Conde, Ana Paula Batista, Gang Zeng, Qianqian Xin, Esper G. Kallas, Jorge Flores, Christian F. Ockenhouse, Christopher Gast Trials (2020-10-15) https://doi.org/ghjkrh
DOI: 10.1186/s13063-020-04775-4 · PMID: 33059771 · PMCID: PMC7558252
984. Role of AS04 in human papillomavirus vaccine: mode of action and clinical profile
Nathalie Garçon, Martine Wettendorff, Marcelle Van Mechelen Expert Opinion on Biological Therapy (2011-04-04) https://doi.org/bvtmpk
DOI: 10.1517/14712598.2011.573624 · PMID: 21457083
985. SARS-CoV-2 spike glycoprotein vaccine candidate NVX-CoV2373 immunogenicity in baboons and protection in mice
Jing-Hui Tian, Nita Patel, Robert Haupt, Haixia Zhou, Stuart Weston, Holly Hammond, James Logue, Alyse D. Portnoff, James Norton, Mimi Guebre-Xabier, … Gale Smith Nature Communications (2021-01-14) https://doi.org/gjh782
DOI: 10.1038/s41467-020-20653-8 · PMID: 33446655 · PMCID: PMC7809486
986. The Coming Age of Insect Cells for Manufacturing and Development of Protein Therapeutics
Christine M. Yee, Andrew J. Zak, Brett D. Hill, Fei Wen Industrial & Engineering Chemistry Research (2018-07-09) https://doi.org/gd332h
DOI: 10.1021/acs.iecr.8b00985 · PMID: 30886455 · PMCID: PMC6420222
987. Matrix-M™ adjuvant enhances immunogenicity of both protein- and modified vaccinia virus Ankara-based influenza vaccines in mice
Sofia E. Magnusson, Arwen F. Altenburg, Karin Lövgren Bengtsson, Fons Bosman, Rory D. de Vries, Guus F. Rimmelzwaan, Linda Stertman Immunologic Research (2018-03-28) https://doi.org/gdd2fw
DOI: 10.1007/s12026-018-8991-x · PMID: 29594879 · PMCID: PMC5899102
988. Immune enhancing properties of the novel Matrix-M™ adjuvant leads to potentiated immune responses to an influenza vaccine in mice
Sofia E. Magnusson, Jenny M. Reimer, Karin H. Karlsson, Lena Lilja, Karin Lövgren Bengtsson, Linda Stertman Vaccine (2013-03) https://doi.org/f2ntg8
DOI: 10.1016/j.vaccine.2013.01.039 · PMID: 23384754
989. Intramuscular Matrix-M-adjuvanted virosomal H5N1 vaccine induces high frequencies of multifunctional Th1 CD4+ cells and strong antibody responses in mice
Abdullah S. Madhun, Lars R. Haaheim, Mona V. Nilsen, Rebecca J. Cox Vaccine (2009-12) https://doi.org/d6cthn
DOI: 10.1016/j.vaccine.2009.09.044 · PMID: 19781678
990. Phase 1–2 Trial of a SARS-CoV-2 Recombinant Spike Protein Nanoparticle Vaccine
Cheryl Keech, Gary Albert, Iksung Cho, Andreana Robertson, Patricia Reed, Susan Neal, Joyce S. Plested, Mingzhu Zhu, Shane Cloney-Clark, Haixia Zhou, … Gregory M. Glenn New England Journal of Medicine (2020-12-10) https://doi.org/gg9q7d
DOI: 10.1056/nejmoa2026920 · PMID: 32877576 · PMCID: PMC7494251
991. Evaluation of a SARS-CoV-2 Vaccine NVX-CoV2373 in Younger and Older Adults
Neil Formica, Raburn Mallory, Gary Albert, Michelle Robinson, Joyce S. Plested, Iksung Cho, Andreana Robertson, Filip Dubovsky, Gregory M. Glenn, for the 2019nCoV-101 Study Group Cold Spring Harbor Laboratory (2021-03-01) https://doi.org/gjh94p
DOI: 10.1101/2021.02.26.21252482
993. Towards an understanding of the adjuvant action of aluminium
Philippa Marrack, Amy S. McKee, Michael W. Munks Nature Reviews Immunology (2009-04) https://doi.org/drcwvf
DOI: 10.1038/nri2510 · PMID: 19247370 · PMCID: PMC3147301
994. DAMP-Inducing Adjuvant and PAMP Adjuvants Parallelly Enhance Protective Type-2 and Type-1 Immune Responses to Influenza Split Vaccination
Tomoya Hayashi, Masatoshi Momota, Etsushi Kuroda, Takato Kusakabe, Shingo Kobari, Kotaro Makisaka, Yoshitaka Ohno, Yusuke Suzuki, Fumika Nakagawa, Michelle S. J. Lee, … Hidetoshi Arima Frontiers in Immunology (2018-11-20) https://doi.org/gfqq89
DOI: 10.3389/fimmu.2018.02619 · PMID: 30515151 · PMCID: PMC6255964
995. Better Adjuvants for Better Vaccines: Progress in Adjuvant Delivery Systems, Modifications, and Adjuvant–Antigen Codelivery
Zhi-Biao Wang, Jing Xu Vaccines (2020-03-13) https://doi.org/gg35vj
DOI: 10.3390/vaccines8010128 · PMID: 32183209 · PMCID: PMC7157724
996. Defining trained immunity and its role in health and disease
Mihai G. Netea, Jorge Domínguez-Andrés, Luis B. Barreiro, Triantafyllos Chavakis, Maziar Divangahi, Elaine Fuchs, Leo A. B. Joosten, Jos W. M. van der Meer, Musa M. Mhlanga, Willem J. M. Mulder, … Eicke Latz Nature Reviews Immunology (2020-03-04) https://doi.org/gg28pr
DOI: 10.1038/s41577-020-0285-6 · PMID: 32132681 · PMCID: PMC7186935
997. Trained Immunity: a Tool for Reducing Susceptibility to and the Severity of SARS-CoV-2 Infection
Mihai G. Netea, Evangelos J. Giamarellos-Bourboulis, Jorge Domínguez-Andrés, Nigel Curtis, Reinout van Crevel, Frank L. van de Veerdonk, Marc Bonten Cell (2020-05) https://doi.org/gg2584
DOI: 10.1016/j.cell.2020.04.042 · PMID: 32437659 · PMCID: PMC7196902
1001. Application of BCG Vaccine for Immune-prophylaxis Among Egyptian Healthcare Workers During the Pandemic of COVID-19
Adel Khattab clinicaltrials.gov (2020-04-17) https://clinicaltrials.gov/ct2/show/NCT04350931
1002. Performance Evaluation of BCG Vaccination in Healthcare Personnel to Reduce the Severity of SARS-COV-2 Infection in Medellín, Colombia, 2020
Universidad de Antioquia clinicaltrials.gov (2020-11-24) https://clinicaltrials.gov/ct2/show/NCT04362124
1005. Reducing Morbidity and Mortality in Health Care Workers Exposed to SARS-CoV-2 by Enhancing Non-specific Immune Responses Through Bacillus Calmette-Guérin Vaccination, a Randomized Controlled Trial
TASK Applied Science clinicaltrials.gov (2020-05-06) https://clinicaltrials.gov/ct2/show/NCT04379336
1006. Randomized Controlled Trial Evaluating the Efficacy of Vaccination With Bacillus Calmette and Guérin (BCG) in the Prevention of COVID-19 Via the Strengthening of Innate Immunity in Health Care Workers
Assistance Publique - Hôpitaux de Paris clinicaltrials.gov (2020-08-17) https://clinicaltrials.gov/ct2/show/NCT04384549
1008. A Randomized Clinical Trial for Enhanced Trained Immune Responses Through Bacillus Calmette-Guérin Vaccination to Prevent Infections by COVID-19: The ACTIVATE II Trial
Hellenic Institute for the Study of Sepsis clinicaltrials.gov (2020-07-10) https://clinicaltrials.gov/ct2/show/NCT04414267
1009. Reducing Hospital Admission of Elderly in SARS-CoV-2 Pandemic Via the Induction of Trained Immunity by Bacillus Calmette-Guérin Vaccination, a Randomized Controlled Trial
Radboud University clinicaltrials.gov (2020-06-03) https://clinicaltrials.gov/ct2/show/NCT04417335
1010. Study to Assess VPM1002 in Reducing Hospital Admissions and/or Severe Respiratory Infectious Diseases in Elderly in COVID-19 Pandemic - Full Text View - ClinicalTrials.govhttps://clinicaltrials.gov/ct2/show/NCT04435379
1015. SARS-CoV-2 and influenza: a comparative overview and treatment implications
Laura D. Manzanares-Meza, Oscar Medina-Contreras Boletín Médico del Hospital Infantil de México (2020-10-23) https://doi.org/gjj2n7
DOI: 10.24875/bmhim.20000183 · PMID: 33064680
1016. Influenza evolution and H3N2 vaccine effectiveness, with application to the 2014/2015 season
Xi Li, Michael W. Deem Protein Engineering Design and Selection (2016-08) https://doi.org/f856v5
DOI: 10.1093/protein/gzw017 · PMID: 27313229 · PMCID: PMC4955871
1017. Neutralizing Activity of BNT162b2-Elicited Serum
Yang Liu, Jianying Liu, Hongjie Xia, Xianwen Zhang, Camila R. Fontes-Garfias, Kena A. Swanson, Hui Cai, Ritu Sarkar, Wei Chen, Mark Cutler, … Pei-Yong Shi New England Journal of Medicine (2021-02-17) https://doi.org/fwsc
DOI: 10.1056/nejmc2102017 · PMCID: PMC7944950
1020. Predicting Influenza H3N2 Vaccine Efficacy From Evolution of the Dominant Epitope
Melia E Bonomo, Michael W Deem Clinical Infectious Diseases (2018-10-01) https://doi.org/gf33js
DOI: 10.1093/cid/ciy323 · PMID: 29672670
1023. An ethical framework for global vaccine allocation
Ezekiel J. Emanuel, Govind Persad, Adam Kern, Allen Buchanan, Cécile Fabre, Daniel Halliday, Joseph Heath, Lisa Herzog, R. J. Leland, Ephrem T. Lemango, … Henry S. Richardson Science (2020-09-11) https://doi.org/ghz7k6
DOI: 10.1126/science.abe2803 · PMID: 32883884
1024. Vaccine optimization for COVID-19: Who to vaccinate first?
Laura Matrajt, Julia Eaton, Tiffany Leung, Elizabeth R. Brown Science Advances (2021-02-03) https://doi.org/ghz7k7
DOI: 10.1126/sciadv.abf1374 · PMID: 33536223
1025. Model-informed COVID-19 vaccine prioritization strategies by age and serostatus
Kate M. Bubar, Kyle Reinholt, Stephen M. Kissler, Marc Lipsitch, Sarah Cobey, Yonatan H. Grad, Daniel B. Larremore Science (2021-02-26) https://doi.org/ght4xk
DOI: 10.1126/science.abe6959 · PMID: 33479118 · PMCID: PMC7963218
1026. Strategic spatiotemporal vaccine distribution increases the survival rate in an infectious disease like Covid-19
Jens Grauer, Hartmut Löwen, Benno Liebchen Scientific Reports (2020-12-09) https://doi.org/ghq7vp
DOI: 10.1038/s41598-020-78447-3 · PMID: 33299029 · PMCID: PMC7726577
1029. Optimal SARS-CoV-2 vaccine allocation using real-time seroprevalence estimates in Rhode Island and Massachusetts
Thu Nguyen-Anh Tran, Nathan Wikle, Joseph Albert, Haider Inam, Emily Strong, Karel Brinda, Scott M Leighow, Fuhan Yang, Sajid Hossain, Justin R Pritchard, … Maciej F Boni Cold Spring Harbor Laboratory (2021-01-15) https://doi.org/gh77b9
DOI: 10.1101/2021.01.12.21249694 · PMID: 33469599 · PMCID: PMC7814845
1039. The Advisory Committee on Immunization Practices’ Interim Recommendation for Use of Pfizer-BioNTech COVID-19 Vaccine — United States, December 2020
Sara E. Oliver, Julia W. Gargano, Mona Marin, Megan Wallace, Kathryn G. Curran, Mary Chamberland, Nancy McClung, Doug Campos-Outcalt, Rebecca L. Morgan, Sarah Mbaeyi, … Kathleen Dooling MMWR. Morbidity and Mortality Weekly Report (2020-12-18) https://doi.org/ghvnsf
DOI: 10.15585/mmwr.mm6950e2 · PMID: 33332292 · PMCID: PMC7745957
1041. The Advisory Committee on Immunization Practices’ Interim Recommendation for Use of Moderna COVID-19 Vaccine — United States, December 2020
Sara E. Oliver, Julia W. Gargano, Mona Marin, Megan Wallace, Kathryn G. Curran, Mary Chamberland, Nancy McClung, Doug Campos-Outcalt, Rebecca L. Morgan, Sarah Mbaeyi, … Kathleen Dooling MMWR. Morbidity and Mortality Weekly Report (2021-01-01) https://doi.org/gh77ch
DOI: 10.15585/mmwr.mm695152e1 · PMID: 33382675
1042. The Advisory Committee on Immunization Practices’ Updated Interim Recommendation for Allocation of COVID-19 Vaccine — United States, December 2020
Kathleen Dooling, Mona Marin, Megan Wallace, Nancy McClung, Mary Chamberland, Grace M. Lee, H. Keipp Talbot, José R. Romero, Beth P. Bell, Sara E. Oliver MMWR. Morbidity and Mortality Weekly Report (2021-01-01) https://doi.org/ghqfvr
DOI: 10.15585/mmwr.mm695152e2 · PMID: 33382671
1045. The Advisory Committee on Immunization Practices’ Interim Recommendation for Use of Janssen COVID-19 Vaccine — United States, February 2021
Sara E. Oliver, Julia W. Gargano, Heather Scobie, Megan Wallace, Stephen C. Hadler, Jessica Leung, Amy E. Blain, Nancy McClung, Doug Campos-Outcalt, Rebecca L. Morgan, … Kathleen Dooling MMWR. Morbidity and Mortality Weekly Report (2021-03-05) https://doi.org/gh77cj
DOI: 10.15585/mmwr.mm7009e4 · PMID: 33661860 · PMCID: PMC7948932
1056. The UK has approved a COVID vaccine — here’s what scientists now want to know
Heidi Ledford, David Cyranoski, Richard Van Noorden Nature (2020-12-03) https://doi.org/gh4xmm
DOI: 10.1038/d41586-020-03441-8 · PMID: 33288887
1093. International Collaboration to Ensure Equitable Access to Vaccines for COVID‐19: The ACT‐Accelerator and the COVAX Facility
MARK ECCLESTON‐TURNER, HARRY UPTON The Milbank Quarterly (2021-03-02) https://doi.org/gh77cc
DOI: 10.1111/1468-0009.12503 · PMID: 33650737
1096. Covax must go beyond proportional allocation of covid vaccines to ensure fair and equitable access
Lisa M Herzog, Ole F Norheim, Ezekiel J Emanuel, Matthew S McCoy BMJ (2021-01-05) https://doi.org/gjgqjv
DOI: 10.1136/bmj.m4853 · PMID: 33402340
1102. Association of Race With Mortality Among Patients Hospitalized With Coronavirus Disease 2019 (COVID-19) at 92 US Hospitals
Baligh R. Yehia, Angela Winegar, Richard Fogel, Mohamad Fakih, Allison Ottenbacher, Christine Jesser, Angelo Bufalino, Ren-Huai Huang, Joseph Cacchione JAMA Network Open (2020-08-18) https://doi.org/ghcspt
DOI: 10.1001/jamanetworkopen.2020.18039 · PMID: 32809033 · PMCID: PMC7435340
1103. Characteristics of and Important Lessons From the Coronavirus Disease 2019 (COVID-19) Outbreak in China
Zunyou Wu, Jennifer M. McGoogan JAMA (2020-04-07) https://doi.org/ggmq43
DOI: 10.1001/jama.2020.2648 · PMID: 32091533
1104. Critical Care Utilization for the COVID-19 Outbreak in Lombardy, Italy
Giacomo Grasselli, Antonio Pesenti, Maurizio Cecconi JAMA (2020-04-28) https://doi.org/ggqf6g
DOI: 10.1001/jama.2020.4031 · PMID: 32167538
1105. Hospitalization Rates and Characteristics of Patients Hospitalized with Laboratory-Confirmed Coronavirus Disease 2019 — COVID-NET, 14 States, March 1–30, 2020
Shikha Garg, Lindsay Kim, Michael Whitaker, Alissa O’Halloran, Charisse Cummings, Rachel Holstein, Mila Prill, Shua J. Chai, Pam D. Kirley, Nisha B. Alden, … Alicia Fry MMWR. Morbidity and Mortality Weekly Report (2020-04-17) https://doi.org/ggsppz
DOI: 10.15585/mmwr.mm6915e3 · PMID: 32298251 · PMCID: PMC7755063
1107. Disparities In Outcomes Among COVID-19 Patients In A Large Health Care System In California
Kristen M. J. Azar, Zijun Shen, Robert J. Romanelli, Stephen H. Lockhart, Kelly Smits, Sarah Robinson, Stephanie Brown, Alice R. Pressman Health Affairs (2020-07-01) https://doi.org/ggx4mf
DOI: 10.1377/hlthaff.2020.00598 · PMID: 32437224
1108. Characteristics Associated with Hospitalization Among Patients with COVID-19 — Metropolitan Atlanta, Georgia, March–April 2020
Marie E. Killerby, Ruth Link-Gelles, Sarah C. Haight, Caroline A. Schrodt, Lucinda England, Danica J. Gomes, Mays Shamout, Kristen Pettrone, Kevin O’Laughlin, Anne Kimball, … CDC COVID-19 Response Clinical Team MMWR. Morbidity and Mortality Weekly Report (2020-06-26) https://doi.org/gg3k6h
DOI: 10.15585/mmwr.mm6925e1 · PMID: 32584797 · PMCID: PMC7316317
1109. Demographic science aids in understanding the spread and fatality rates of COVID-19
Jennifer Beam Dowd, Liliana Andriano, David M. Brazel, Valentina Rotondi, Per Block, Xuejie Ding, Yan Liu, Melinda C. Mills Proceedings of the National Academy of Sciences (2020-05-05) https://doi.org/ggsd5b
DOI: 10.1073/pnas.2004911117 · PMID: 32300018 · PMCID: PMC7211934
1110. ‐19 and Older Adults: What We Know
Zainab Shahid, Ricci Kalayanamitra, Brendan McClafferty, Douglas Kepko, Devyani Ramgobin, Ravi Patel, Chander Shekher Aggarwal, Ramarao Vunnam, Nitasa Sahu, Dhirisha Bhatt, … Rohit Jain Journal of the American Geriatrics Society (2020-04-20) https://doi.org/ggxgsb
DOI: 10.1111/jgs.16472 · PMID: 32255507 · PMCID: PMC7262251
1111. Features of 20 133 UK patients in hospital with covid-19 using the ISARIC WHO Clinical Characterisation Protocol: prospective observational cohort study
Annemarie B Docherty, Ewen M Harrison, Christopher A Green, Hayley E Hardwick, Riinu Pius, Lisa Norman, Karl A Holden, Jonathan M Read, Frank Dondelinger, Gail Carson, … Malcolm G Semple BMJ (2020-05-22) https://doi.org/ggw4nh
DOI: 10.1136/bmj.m1985 · PMID: 32444460 · PMCID: PMC7243036
1112. Impact of sex and gender on COVID-19 outcomes in Europe
Catherine Gebhard, Vera Regitz-Zagrosek, Hannelore K. Neuhauser, Rosemary Morgan, Sabra L. Klein Biology of Sex Differences (2020-05-25) https://doi.org/ghbvck
DOI: 10.1186/s13293-020-00304-9 · PMID: 32450906 · PMCID: PMC7247289
1115. Sex-specific clinical characteristics and prognosis of coronavirus disease-19 infection in Wuhan, China: A retrospective study of 168 severe patients
Yifan Meng, Ping Wu, Wanrong Lu, Kui Liu, Ke Ma, Liang Huang, Jiaojiao Cai, Hong Zhang, Yu Qin, Haiying Sun, … Peng Wu PLOS Pathogens (2020-04-28) https://doi.org/ggv3zn
DOI: 10.1371/journal.ppat.1008520 · PMID: 32343745 · PMCID: PMC7209966
1116. Sex differences in renal angiotensin converting enzyme 2 (ACE2) activity are 17β-oestradiol-dependent and sex chromosome-independent
Jun Liu, Hong Ji, Wei Zheng, Xie Wu, Janet J Zhu, Arthur P Arnold, Kathryn Sandberg Biology of Sex Differences (2010) https://doi.org/bbx3r6
DOI: 10.1186/2042-6410-1-6 · PMID: 21208466 · PMCID: PMC3010099
1123. Severe obesity, increasing age and male sex are independently associated with worse in-hospital outcomes, and higher in-hospital mortality, in a cohort of patients with COVID-19 in the Bronx, New York
Leonidas Palaiodimos, Damianos G. Kokkinidis, Weijia Li, Dimitrios Karamanis, Jennifer Ognibene, Shitij Arora, William N. Southern, Christos S. Mantzoros Metabolism (2020-07) https://doi.org/ggx229
DOI: 10.1016/j.metabol.2020.154262 · PMID: 32422233 · PMCID: PMC7228874
1124. Features of 16,749 hospitalised UK patients with COVID-19 using the ISARIC WHO Clinical Characterisation Protocol
AB Docherty, EM Harrison, CA Green, H Hardwick, R Pius, L Norman, KA Holden, JM Read, F Dondelinger, G Carson, … ISARIC4C Investigators Cold Spring Harbor Laboratory (2020-04-28) https://doi.org/ggtdtb
DOI: 10.1101/2020.04.23.20076042
1126. Besides population age structure, health and other demographic factors can contribute to understanding the COVID-19 burden
Marília R. Nepomuceno, Enrique Acosta, Diego Alburez-Gutierrez, José Manuel Aburto, Alain Gagnon, Cássio M. Turra Proceedings of the National Academy of Sciences (2020-06-23) https://doi.org/gg33qx
DOI: 10.1073/pnas.2008760117 · PMID: 32576710 · PMCID: PMC7322063
1128. The SOFA (Sepsis-related Organ Failure Assessment) score to describe organ dysfunction/failure
J. -L. Vincent, R. Moreno, J. Takala, S. Willatts, A. De Mendonça, H. Bruining, C. K. Reinhart, P. M. Suter, L. G. Thijs Intensive Care Medicine (1996-07) https://doi.org/bpkxdw
DOI: 10.1007/bf01709751 · PMID: 8844239
1129. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3)
Mervyn Singer, Clifford S. Deutschman, Christopher Warren Seymour, Manu Shankar-Hari, Djillali Annane, Michael Bauer, Rinaldo Bellomo, Gordon R. Bernard, Jean-Daniel Chiche, Craig M. Coopersmith, … Derek C. Angus JAMA (2016-02-23) https://doi.org/gdrcdh
DOI: 10.1001/jama.2016.0287 · PMID: 26903338 · PMCID: PMC4968574
1133. Susceptibility of Southwestern American Indian Tribes to Coronavirus Disease 2019 (COVID‐19)
Monika Kakol, Dona Upson, Akshay Sood The Journal of Rural Health (2020-06) https://doi.org/ggtzkq
DOI: 10.1111/jrh.12451 · PMID: 32304251 · PMCID: PMC7264672
1135. Addressing inequities in COVID-19 morbidity and mortality: research and policy recommendations
Monica L Wang, Pamela Behrman, Akilah Dulin, Monica L Baskin, Joanna Buscemi, Kassandra I Alcaraz, Carly M Goldstein, Tiffany L Carson, Megan Shen, Marian Fitzgibbon Translational Behavioral Medicine (2020-06) https://doi.org/gg3389
DOI: 10.1093/tbm/ibaa055 · PMID: 32542349 · PMCID: PMC7337775
1136. Historical Environmental Racism, Structural Inequalities, and Dik’os Ntsaaígíí-19 (COVID-19) on Navajo Nation
Nicholet A. Deschine Parkhurst, Kimberly R. Huyser, Aggie J. Yellow Horse Journal of Indigenous Social Development (2020-11-02) https://journalhosting.ucalgary.ca/index.php/jisd/article/view/70753
1138. Factors associated with COVID-19-related death using OpenSAFELY
Elizabeth J. Williamson, Alex J. Walker, Krishnan Bhaskaran, Seb Bacon, Chris Bates, Caroline E. Morton, Helen J. Curtis, Amir Mehrkar, David Evans, Peter Inglesby, … Ben Goldacre Nature (2020-07-08) https://doi.org/gg39n7
DOI: 10.1038/s41586-020-2521-4 · PMID: 32640463
1139. Implications of biogeography of human populations for “race” and medicine
Sarah A Tishkoff, Kenneth K Kidd Nature Genetics (2004-10-26) https://doi.org/d2xq92
DOI: 10.1038/ng1438 · PMID: 15507999
1140. African Genetic Diversity: Implications for Human Demographic History, Modern Human Origins, and Complex Disease Mapping
Michael C. Campbell, Sarah A. Tishkoff Annual Review of Genomics and Human Genetics (2008-09) https://doi.org/cphggp
DOI: 10.1146/annurev.genom.9.081307.164258 · PMID: 18593304 · PMCID: PMC2953791
1141. NIH must confront the use of race in science
Michael Yudell, Dorothy Roberts, Rob DeSalle, Sarah Tishkoff, 70 signatories Science (2020-09-10) https://doi.org/ghcm7s
DOI: 10.1126/science.abd4842 · PMID: 32913094
1142. Interferon-Induced Transmembrane Protein 3 Genetic Variant rs12252-C Associated With Disease Severity in Coronavirus Disease 2019
Yonghong Zhang, Ling Qin, Yan Zhao, Ping Zhang, Bin Xu, Kang Li, Lianchun Liang, Chi Zhang, Yanchao Dai, Yingmei Feng, … Ronghua Jin The Journal of Infectious Diseases (2020-07-01) https://doi.org/ggv3tj
DOI: 10.1093/infdis/jiaa224 · PMID: 32348495 · PMCID: PMC7197559
1144. APOE e4 Genotype Predicts Severe COVID-19 in the UK Biobank Community Cohort
Chia-Ling Kuo, Luke C Pilling, Janice L Atkins, Jane AH Masoli, João Delgado, George A Kuchel, David Melzer The Journals of Gerontology: Series A (2020-11) https://doi.org/ggx4ng
DOI: 10.1093/gerona/glaa131 · PMID: 32451547 · PMCID: PMC7314139
1145. Genome-wide CRISPR screen reveals host genes that regulate SARS-CoV-2 infection
Jin Wei, Mia Madel Alfajaro, Ruth E. Hanna, Peter C. DeWeirdt, Madison S. Strine, William J. Lu-Culligan, Shang-Min Zhang, Vincent R. Graziano, Cameron O. Schmitz, Jennifer S. Chen, … Craig B. Wilen Cold Spring Harbor Laboratory (2020-06-17) https://doi.org/dzz3
DOI: 10.1101/2020.06.16.155101 · PMID: 32869025 · PMCID: PMC7457610
1146. New insights into genetic susceptibility of COVID-19: an ACE2 and TMPRSS2 polymorphism analysis
Yuan Hou, Junfei Zhao, William Martin, Asha Kallianpur, Mina K. Chung, Lara Jehi, Nima Sharifi, Serpil Erzurum, Charis Eng, Feixiong Cheng BMC Medicine (2020-07-15) https://doi.org/gg445n
DOI: 10.1186/s12916-020-01673-z · PMID: 32664879 · PMCID: PMC7360473
1147. Seroprevalence of anti-SARS-CoV-2 IgG antibodies in Kenyan blood donors
Sophie Uyoga, Ifedayo M.O. Adetifa, Henry K. Karanja, James Nyagwange, James Tuju, Perpetual Wanjiku, Rashid Aman, Mercy Mwangangi, Patrick Amoth, Kadondi Kasera, … George M. Warimwe Cold Spring Harbor Laboratory (2020-07-29) https://doi.org/ghcm7p
DOI: 10.1101/2020.07.27.20162693
1148. High SARS-CoV-2 seroprevalence in Health Care Workers but relatively low numbers of deaths in urban Malawi
Marah G. Chibwana, Khuzwayo C. Jere, Raphael Kamn’gona, Jonathan Mandolo, Vincent Katunga-Phiri, Dumizulu Tembo, Ndaona Mitole, Samantha Musasa, Simon Sichone, Agness Lakudzala, … Kondwani C. Jambo Cold Spring Harbor Laboratory (2020-08-01) https://doi.org/ghcm7q
DOI: 10.1101/2020.07.30.20164970 · PMID: 32766597 · PMCID: PMC7402052
1151. Quantifying the social distancing privilege gap: a longitudinal study of smartphone movement
Nabarun Dasgupta, Michele Jonsson Funk, Allison Lazard, Benjamin Eugene White, Stephen W. Marshall Cold Spring Harbor Laboratory (2020-05-08) https://doi.org/gg79qk
DOI: 10.1101/2020.05.03.20084624
1152. Uncovering socioeconomic gaps in mobility reduction during the COVID-19 pandemic using location data
Samuel P. Fraiberger, Pablo Astudillo, Lorenzo Candeago, Alex Chunet, Nicholas K. W. Jones, Maham Faisal Khan, Bruno Lepri, Nancy Lozano Gracia, Lorenzo Lucchini, Emanuele Massaro, Aleister Montfort arXiv (2020-07-28) https://arxiv.org/abs/2006.15195
1153. Mobility network models of COVID-19 explain inequities and inform reopening
Serina Chang, Emma Pierson, Pang Wei Koh, Jaline Gerardin, Beth Redbird, David Grusky, Jure Leskovec Nature (2020-11-10) https://doi.org/ghjmt2
DOI: 10.1038/s41586-020-2923-3 · PMID: 33171481
1155. Differential occupational risk for COVID‐19 and other infection exposure according to race and ethnicity
Devan Hawkins American Journal of Industrial Medicine (2020-06-15) https://doi.org/gg3rb2
DOI: 10.1002/ajim.23145 · PMID: 32539166 · PMCID: PMC7323065
1156. Estimating the burden of United States workers exposed to infection or disease: A key factor in containing risk of COVID-19 infection
Marissa G. Baker, Trevor K. Peckham, Noah S. Seixas PLOS ONE (2020-04-28) https://doi.org/ggtx7c
DOI: 10.1371/journal.pone.0232452 · PMID: 32343747 · PMCID: PMC7188235
1162. Racial Disparity in COVID-19 Deaths: Seeking Economic Roots with Census data.
John McLaren National Bureau of Economic Research (2020-06-22) https://www.nber.org/papers/w27407
1163. Mortality, Admissions, and Patient Census at SNFs in 3 US Cities During the COVID-19 Pandemic
Michael L. Barnett, Lissy Hu, Thomas Martin, David C. Grabowski JAMA (2020-08-04) https://doi.org/gg3387
DOI: 10.1001/jama.2020.11642 · PMID: 32579161 · PMCID: PMC7315390
1167. Under One Roof: A Review of Research on Intergenerational Coresidence and Multigenerational Households in the United States
Jennifer Reid Keene, Christie D. Batson Sociology Compass (2010-08) https://doi.org/fsrcr7
DOI: 10.1111/j.1751-9020.2010.00306.x
1168. Chaos and the macrosetting: The role of poverty and socioeconomic status.
Gary W. Evans, John Eckenrode, Lyscha A. Marcynyszyn American Psychological Association (APA) (2010-01-12) https://doi.org/c76n3g
DOI: 10.1037/12057-014
1173. Non-communicable disease syndemics: poverty, depression, and diabetes among low-income populations
Emily Mendenhall, Brandon A Kohrt, Shane A Norris, David Ndetei, Dorairaj Prabhakaran The Lancet (2017-03) https://doi.org/gddg84
DOI: 10.1016/s0140-6736(17)30402-6 · PMID: 28271846 · PMCID: PMC5491333
1174. Obesity and poverty paradox in developed countries
Wioletta Żukiewicz-Sobczak, Paula Wróblewska, Jacek Zwoliński, Jolanta Chmielewska-Badora, Piotr Adamczuk, Ewelina Krasowska, Jerzy Zagórski, Anna Oniszczuk, Jacek Piątek, Wojciech Silny Annals of Agricultural and Environmental Medicine (2014-09-04) https://doi.org/f6jhzc
DOI: 10.5604/12321966.1120608 · PMID: 25292135
1176. Fast Food Patronage and Obesity Prevalence During the COVID‐19 Pandemic: An Alternative Explanation
Candice A. Myers, Stephanie T. Broyles Obesity (2020-09-03) https://doi.org/gg6v84
DOI: 10.1002/oby.22993 · PMID: 32741130 · PMCID: PMC7435526
1177. The global food syndemic: The impact of food insecurity, Malnutrition and obesity on the healthspan amid the COVID-19 pandemic
Martha I. Huizar, Ross Arena, Deepika R. Laddu Progress in Cardiovascular Diseases (2020-07) https://doi.org/gg4r3h
DOI: 10.1016/j.pcad.2020.07.002 · PMID: 32653438 · PMCID: PMC7347484
1178. Stress, chronic inflammation, and emotional and physical well-being: Concurrent effects and chronic sequelae
George P. Chrousos Journal of Allergy and Clinical Immunology (2000-11) https://doi.org/bgx7hn
DOI: 10.1067/mai.2000.110163 · PMID: 11080744
1179. Chronic psychological stress and the regulation of pro-inflammatory cytokines: A glucocorticoid-resistance model.
Gregory E. Miller, Sheldon Cohen, A. Kim Ritchey Health Psychology (2002) https://doi.org/dj5r8b
DOI: 10.1037/0278-6133.21.6.531
1180. Chronic stress, daily stressors, and circulating inflammatory markers.
Jean-Philippe Gouin, Ronald Glaser, William B. Malarkey, David Beversdorf, Janice Kiecolt-Glaser Health Psychology (2012-03) https://doi.org/dkz9tr
DOI: 10.1037/a0025536 · PMID: 21928900 · PMCID: PMC3253267
1183. Exposure to air pollution and COVID-19 mortality in the United States: A nationwide cross-sectional study
Xiao Wu, Rachel C Nethery, M Benjamin Sabath, Danielle Braun, Francesca Dominici Cold Spring Harbor Laboratory (2020-04-27) https://doi.org/ggrpcj
DOI: 10.1101/2020.04.05.20054502 · PMID: 32511651 · PMCID: PMC7277007
1184. Global estimates of mortality associated with long-term exposure to outdoor fine particulate matter
Richard Burnett, Hong Chen, Mieczysław Szyszkowicz, Neal Fann, Bryan Hubbell, C. Arden Pope, Joshua S. Apte, Michael Brauer, Aaron Cohen, Scott Weichenthal, … Joseph V. Spadaro Proceedings of the National Academy of Sciences (2018-09-18) https://doi.org/gfgbcx
DOI: 10.1073/pnas.1803222115 · PMID: 30181279 · PMCID: PMC6156628
1185. Early-Life Air Pollution Exposure, Neighborhood Poverty, and Childhood Asthma in the United States, 1990–2014
Nicole Kravitz-Wirtz, Samantha Teixeira, Anjum Hajat, Bongki Woo, Kyle Crowder, David Takeuchi International Journal of Environmental Research and Public Health (2018-05-30) https://doi.org/gdvwp9
DOI: 10.3390/ijerph15061114 · PMID: 29848979 · PMCID: PMC6025399
1186. Early life stress, air pollution, inflammation, and disease: An integrative review and immunologic model of social-environmental adversity and lifespan health
Hector A. Olvera Alvarez, Laura D. Kubzansky, Matthew J. Campen, George M. Slavich Neuroscience & Biobehavioral Reviews (2018-09) https://doi.org/gd46bm
DOI: 10.1016/j.neubiorev.2018.06.002 · PMID: 29874545 · PMCID: PMC6082389
1187. Covid-19 and Disparities in Nutrition and Obesity
Matthew J. Belanger, Michael A. Hill, Angeliki M. Angelidi, Maria Dalamaga, James R. Sowers, Christos S. Mantzoros New England Journal of Medicine (2020-09-10) https://doi.org/gg475x
DOI: 10.1056/nejmp2021264 · PMID: 32668105
1188. Systemic racism, chronic health inequities, and ‐19: A syndemic in the making?
Clarence C. Gravlee American Journal of Human Biology (2020-08-04) https://doi.org/ghcxwk
DOI: 10.1002/ajhb.23482 · PMID: 32754945 · PMCID: PMC7441277
1190. Coronavirus Disease (COVID-19): A primer for emergency physicians
Summer Chavez, Brit Long, Alex Koyfman, Stephen Y. Liang The American Journal of Emergency Medicine (2020-03) https://doi.org/ggr22z
DOI: 10.1016/j.ajem.2020.03.036 · PMID: 32265065 · PMCID: PMC7102516
1194. FAQs for COVID-19 Claims Reimbursement to Health Care Providers and Facilities for Testing, Treatment and Vaccine Administration
Official web site of the U.S. Health Resources & Services Administration https://www.hrsa.gov/coviduninsuredclaim/frequently-asked-questions
1195. Potential association between COVID-19 mortality and health-care resource availability
Yunpeng Ji, Zhongren Ma, Maikel P Peppelenbosch, Qiuwei Pan The Lancet Global Health (2020-04) https://doi.org/ggqscd
DOI: 10.1016/s2214-109x(20)30068-1 · PMID: 32109372 · PMCID: PMC7128131
1198. An Ethical Dilemma in SARS-Cov-2 Pandemic : Who Gets the Ventilator?
Dumache Raluca, Ciocan Veronica, Muresan Camelia Oana, Enache Alexandra European Scientific Journal ESJ (2020-07-31) https://doi.org/ghfprk
DOI: 10.19044/esj.2020.v16n21p24
1199. Planning Hospital Needs for Ventilators and Respiratory Therapists in the COVID-19 Crisis
John Raffensperger, Marygail Brauner, R. Briggs Rand Corporation (2020) https://doi.org/ghfprp
DOI: 10.7249/pea228-1
1200. Fair Allocation of Vaccines, Ventilators and Antiviral Treatments: Leaving No Ethical Value Behind in Health Care Rationing
Parag A. Pathak, Tayfun Sönmez, M. Utku Ünver, M. Bumin Yenmez arXiv (2021-01-21) https://arxiv.org/abs/2008.00374
1201. Reallocating ventilators during the coronavirus disease 2019 pandemic: Is it ethical?
Quyen Chu, Ricardo Correa, Tracey L. Henry, Kyle A. McGregor, Hanni Stoklosa, Loren Robinson, Sachin Jha, Alagappan Annamalai, Benson S. Hsu, Rohit Gupta, … SreyRam Kuy Surgery (2020-09) https://doi.org/ghfprb
DOI: 10.1016/j.surg.2020.04.044 · PMID: 32616345 · PMCID: PMC7205622
1202. Ethics Lessons From Seattle’s Early Experience With COVID-19
Denise M. Dudzinski, Benjamin Y. Hoisington, Crystal E. Brown The American Journal of Bioethics (2020-06-18) https://doi.org/ghfprc
DOI: 10.1080/15265161.2020.1764137 · PMID: 32552455
1203. Rationing Limited Healthcare Resources in the COVID‐19 Era and Beyond: Ethical Considerations Regarding Older Adults
Timothy W. Farrell, Leslie Francis, Teneille Brown, Lauren E. Ferrante, Eric Widera, Ramona Rhodes, Tony Rosen, Ula Hwang, Leah J. Witt, Niranjan Thothala, … Debra Saliba Journal of the American Geriatrics Society (2020-06-14) https://doi.org/ggvt7z
DOI: 10.1111/jgs.16539 · PMID: 32374466 · PMCID: PMC7267288
1204. Paediatric ethical issues during the COVID‐19 pandemic are not just about ventilator triage
Marlyse F. Haward, Gregory P. Moore, John Lantos, Annie Janvier Acta Paediatrica (2020-05-20) https://doi.org/ggv24n
DOI: 10.1111/apa.15334 · PMID: 32364256 · PMCID: PMC7267437
1205. Ethical Challenges Arising in the COVID-19 Pandemic: An Overview from the Association of Bioethics Program Directors (ABPD) Task Force
Amy L. McGuire, Mark P. Aulisio, F. Daniel Davis, Cheryl Erwin, Thomas D. Harter, Reshma Jagsi, Robert Klitzman, Robert Macauley, Eric Racine, Susan M. Wolf, … The COVID-19 Task Force of the Association of Bioethics Program Directors (ABPD) The American Journal of Bioethics (2020-06-08) https://doi.org/gg6c5k
DOI: 10.1080/15265161.2020.1764138 · PMID: 32511078
1206. Disability, Ethics, and Health Care in the COVID-19 Pandemic
Maya Sabatello, Teresa Blankmeyer Burke, Katherine E. McDonald, Paul S. Appelbaum American Journal of Public Health (2020-10) https://doi.org/ghfprm
DOI: 10.2105/ajph.2020.305837 · PMID: 32816541 · PMCID: PMC7483109
1208. Colorblind Algorithms: Racism in the Era of COVID-19
J. Corey Williams, Nientara Anderson, Myra Mathis, Ezelle Sanford, Jeffrey Eugene, Jessica Isom Journal of the National Medical Association (2020-10) https://doi.org/ghfpq8
DOI: 10.1016/j.jnma.2020.05.010 · PMID: 32563687
1209. Structural Racism, Social Risk Factors, and Covid-19 — A Dangerous Convergence for Black Americans
Leonard E. Egede, Rebekah J. Walker New England Journal of Medicine (2020-09-17) https://doi.org/gg56nc
DOI: 10.1056/nejmp2023616 · PMID: 32706952 · PMCID: PMC7747672
1212. Ethical Dilemmas in Covid-19 Medical Care: Is a Problematic Triage Protocol Better or Worse than No Protocol at All?
Sheri Fink The American Journal of Bioethics (2020-07-27) https://doi.org/gg6nqn
DOI: 10.1080/15265161.2020.1788663 · PMID: 32716771
1213. Developing an Ethics Framework for Allocating Remdesivir in the COVID-19 Pandemic
Sarah Lim, Debra A. DeBruin, Jonathon P. Leider, Nneka Sederstrom, Ruth Lynfield, Jason V. Baker, Susan Kline, Sarah Kesler, Stacey Rizza, Joel Wu, … Susan M. Wolf Mayo Clinic Proceedings (2020-09) https://doi.org/ghfpq9
DOI: 10.1016/j.mayocp.2020.06.016 · PMID: 32861338 · PMCID: PMC7305893
1214. Ethically Allocating COVID-19 Drugs Via Pre-approval Access and Emergency Use Authorization
Jamie Webb, Lesha D. Shah, Holly Fernandez Lynch The American Journal of Bioethics (2020-08-25) https://doi.org/ghfprd
DOI: 10.1080/15265161.2020.1795529
1216. Adopting an Anti-Racist Model of COVID-19 Drug Allocation and Prioritization
Akilah A. Jefferson The American Journal of Bioethics (2020-08-25) https://doi.org/ghggz5
DOI: 10.1080/15265161.2020.1795541
1219. The Equitable Distribution of COVID-19 Therapeutics and Vaccines
Thomas J. Bollyky, Lawrence O. Gostin, Margaret A. Hamburg JAMA (2020-06-23) https://doi.org/ggvcvt
DOI: 10.1001/jama.2020.6641 · PMID: 32379268
1220. Recruitment and participation in clinical trials: Socio-demographic, rural/urban, and health care access predictors
Claudia R. Baquet, Patricia Commiskey, C. Daniel Mullins, Shiraz I. Mishra Cancer Detection and Prevention (2006-01) https://doi.org/bk2k4g
DOI: 10.1016/j.cdp.2005.12.001 · PMID: 16495020 · PMCID: PMC3276312
1222. Racial/ethnic differences in clinical trial enrollment, refusal rates, ineligibility, and reasons for decline among patients at sites in the National Cancer Institute’s Community Cancer Centers Program
Aisha T. Langford, Ken Resnicow, Eileen P. Dimond, Andrea M. Denicoff, Diane St. Germain, Worta McCaskill-Stevens, Rebecca A. Enos, Angela Carrigan, Kathy Wilkinson, Ronald S. Go Cancer (2014-03-15) https://doi.org/ghgg2p
DOI: 10.1002/cncr.28483 · PMID: 24327389 · PMCID: PMC3947654
1224. Participation in Surgical Oncology Clinical Trials: Gender-, Race/Ethnicity-, and Age-based Disparities
John H. Stewart, Alain G. Bertoni, Jennifer L. Staten, Edward A. Levine, Cary P. Gross Annals of Surgical Oncology (2007-08-08) https://doi.org/cq7wqs
DOI: 10.1245/s10434-007-9500-y · PMID: 17682824
1225. Inclusion, Analysis, and Reporting of Sex and Race/Ethnicity in Clinical Trials: Have We Made Progress?
Stacie E. Geller, Abby Koch, Beth Pellettieri, Molly Carnes Journal of Women’s Health (2011-03) https://doi.org/dhzxk7
DOI: 10.1089/jwh.2010.2469 · PMID: 21351877 · PMCID: PMC3058895
1226. The Representation of Gender and Race/Ethnic Groups in Randomized Clinical Trials of Individuals with Systemic Lupus Erythematosus
Titilola Falasinnu, Yashaar Chaichian, Michelle B. Bass, Julia F. Simard Current Rheumatology Reports (2018-03-17) https://doi.org/ghjmpz
DOI: 10.1007/s11926-018-0728-2 · PMID: 29550947 · PMCID: PMC5857270
1227. Racial Disproportionality in Covid Clinical Trials
Daniel B. Chastain, Sharmon P. Osae, Andrés F. Henao-Martínez, Carlos Franco-Paredes, Joeanna S. Chastain, Henry N. Young New England Journal of Medicine (2020-08-27) https://doi.org/gg7vcf
DOI: 10.1056/nejmp2021971 · PMID: 32780573
1228. Othering and Being Othered in the Context of Health Care Services
Joy L. Johnson, Joan L. Bottorff, Annette J. Browne, Sukhdev Grewal, B. Ann Hilton, Heather Clarke Health Communication (2004-04) https://doi.org/cvxqm4
DOI: 10.1207/s15327027hc1602_7 · PMID: 15090288
1229. A decade of studying implicit racial/ethnic bias in healthcare providers using the implicit association test
Ivy W. Maina, Tanisha D. Belton, Sara Ginzberg, Ajit Singh, Tiffani J. Johnson Social Science & Medicine (2018-02) https://doi.org/gdfwd9
DOI: 10.1016/j.socscimed.2017.05.009 · PMID: 28532892
1230. A Systematic Review of the Impact of Physician Implicit Racial Bias on Clinical Decision Making
Erin Dehon, Nicole Weiss, Jonathan Jones, Whitney Faulconer, Elizabeth Hinton, Sarah Sterling Academic Emergency Medicine (2017-08) https://doi.org/gbw5pk
DOI: 10.1111/acem.13214 · PMID: 28472533
1231. Aversive racism and medical interactions with Black patients: A field study
Louis A. Penner, John F. Dovidio, Tessa V. West, Samuel L. Gaertner, Terrance L. Albrecht, Rhonda K. Dailey, Tsveti Markova Journal of Experimental Social Psychology (2010-03) https://doi.org/dc5342
DOI: 10.1016/j.jesp.2009.11.004 · PMID: 20228874 · PMCID: PMC2835170
1233. A Systematic Review Of The Food And Drug Administration’s “Exception From Informed Consent” Pathway
William B. Feldman, Spencer Phillips Hey, Aaron S. Kesselheim Health Affairs (2018-10) https://doi.org/ghjmqw
DOI: 10.1377/hlthaff.2018.0501 · PMID: 30273035
1234. The legacy of the tuskegee syphilis experiments for emergency exception from informed consent
Terri A. Schmidt Annals of Emergency Medicine (2003-01) https://doi.org/fw3kvs
DOI: 10.1067/mem.2003.17 · PMID: 12514686
1236. Racial Differences in T-Lymphocyte Response to Glucocorticoids
Monica J. Federico, Ronina A. Covar, Eleanor E. Brown, Donald Y. M. Leung, Joseph D. Spahn Chest (2005-02) https://doi.org/bjfcf6
DOI: 10.1378/chest.127.2.571 · PMID: 15705998
1238. Chronic stress, glucocorticoid receptor resistance, inflammation, and disease risk
S. Cohen, D. Janicki-Deverts, W. J. Doyle, G. E. Miller, E. Frank, B. S. Rabin, R. B. Turner Proceedings of the National Academy of Sciences (2012-04-02) https://doi.org/f4n6r8
DOI: 10.1073/pnas.1118355109 · PMID: 22474371 · PMCID: PMC3341031
1239. Opportunities and obstacles for deep learning in biology and medicine
Travers Ching, Daniel S. Himmelstein, Brett K. Beaulieu-Jones, Alexandr A. Kalinin, Brian T. Do, Gregory P. Way, Enrico Ferrero, Paul-Michael Agapow, Michael Zietz, Michael M. Hoffman, … Casey S. Greene Journal of The Royal Society Interface (2018-04-04) https://doi.org/gddkhn
DOI: 10.1098/rsif.2017.0387 · PMID: 29618526 · PMCID: PMC5938574
1240. Opportunities and obstacles for deep learning in biology and medicine [update in progress]
Casey S. Greene, Daniel S. Himmelstein, Daniel C. Elton, Brock C. Christensen, Anthony Gitter, Alexander J. Titus, Joshua J. Levy Manubot (2020-11-10) https://greenelab.github.io/deep-review/
1245. SARS-CoV-2 and COVID-19: An Evolving Review of Diagnostics and Therapeutics
Halie M. Rando, Casey S. Greene, Michael P. Robson, Simina M. Boca, Nils Wellhausen, Ronan Lordan, Christian Brueffer, Sandipan Ray, Lucy D\’Agostino McGowan, Anthony Gitter, … Rishi Raj Goel Manubot (2021-02-08) https://greenelab.github.io/covid19-review/
1248. Potent binding of 2019 novel coronavirus spike protein by a SARS coronavirus-specific human monoclonal antibody
Xiaolong Tian, Cheng Li, Ailing Huang, Shuai Xia, Sicong Lu, Zhengli Shi, Lu Lu, Shibo Jiang, Zhenlin Yang, Yanling Wu, Tianlei Ying Cold Spring Harbor Laboratory (2020-01-28) https://doi.org/ggjqfd
DOI: 10.1101/2020.01.28.923011
1249. Integrative Bioinformatics Analysis Provides Insight into the Molecular Mechanisms of 2019-nCoV
Xiang He, Lei Zhang, Qin Ran, Junyi Wang, Anying Xiong, Dehong Wu, Feng Chen, Guoping Li Cold Spring Harbor Laboratory (2020-02-05) https://doi.org/ggrbd8
DOI: 10.1101/2020.02.03.20020206
1250. Diarrhea may be underestimated: a missing link in 2019 novel coronavirus
Weicheng Liang, Zhijie Feng, Shitao Rao, Cuicui Xiao, Ze-Xiao Lin, Qi Zhang, Qi Wei Cold Spring Harbor Laboratory (2020-02-17) https://doi.org/ggrbdw
DOI: 10.1101/2020.02.03.20020289
1251. Specific ACE2 Expression in Cholangiocytes May Cause Liver Damage After 2019-nCoV Infection
Xiaoqiang Chai, Longfei Hu, Yan Zhang, Weiyu Han, Zhou Lu, Aiwu Ke, Jian Zhou, Guoming Shi, Nan Fang, Jia Fan, … Fei Lan Cold Spring Harbor Laboratory (2020-02-04) https://doi.org/ggq626
DOI: 10.1101/2020.02.03.931766
1252. Recapitulation of SARS-CoV-2 Infection and Cholangiocyte Damage with Human Liver Organoids
Bing Zhao, Chao Ni, Ran Gao, Yuyan Wang, Li Yang, Jinsong Wei, Ting Lv, Jianqing Liang, Qisheng Zhang, Wei Xu, … Xinhua Lin Cold Spring Harbor Laboratory (2020-03-17) https://doi.org/ggq648
DOI: 10.1101/2020.03.16.990317
1253. ACE2 expression by colonic epithelial cells is associated with viral infection, immunity and energy metabolism
Jun Wang, Shanmeizi Zhao, Ming Liu, Zhiyao Zhao, Yiping Xu, Ping Wang, Meng Lin, Yanhui Xu, Bing Huang, Xiaoyu Zuo, … Yuxia Zhang Cold Spring Harbor Laboratory (2020-02-07) https://doi.org/ggrfbx
DOI: 10.1101/2020.02.05.20020545
1254. The Pathogenicity of SARS-CoV-2 in hACE2 Transgenic Mice
Linlin Bao, Wei Deng, Baoying Huang, Hong Gao, Jiangning Liu, Lili Ren, Qiang Wei, Pin Yu, Yanfeng Xu, Feifei Qi, … Chuan Qin Cold Spring Harbor Laboratory (2020-02-28) https://doi.org/dph2
DOI: 10.1101/2020.02.07.939389
1255. Caution on Kidney Dysfunctions of COVID-19 Patients
Zhen Li, Ming Wu, Jiwei Yao, Jie Guo, Xiang Liao, Siji Song, Jiali Li, Guangjie Duan, Yuanxiu Zhou, Xiaojun Wu, … Junan Yan Cold Spring Harbor Laboratory (2020-03-27) https://doi.org/ggq627
DOI: 10.1101/2020.02.08.20021212
1256. Acute renal impairment in coronavirus-associated severe acute respiratory syndrome
Kwok Hong Chu, Wai Kay Tsang, Colin S. Tang, Man Fai Lam, Fernand M. Lai, Ka Fai To, Ka Shun Fung, Hon Lok Tang, Wing Wa Yan, Hilda W. H. Chan, … Kar Neng Lai Kidney International (2005-02) https://doi.org/b7tgtx
DOI: 10.1111/j.1523-1755.2005.67130.x · PMID: 15673319 · PMCID: PMC7112337
1257. Single-cell Analysis of ACE2 Expression in Human Kidneys and Bladders Reveals a Potential Route of 2019-nCoV Infection
Wei Lin, Longfei Hu, Yan Zhang, Joshua D. Ooi, Ting Meng, Peng Jin, Xiang Ding, Longkai Peng, Lei Song, Zhou Xiao, … Yong Zhong Cold Spring Harbor Laboratory (2020-02-18) https://doi.org/ggq629
DOI: 10.1101/2020.02.08.939892
1258. The immune vulnerability landscape of the 2019 Novel Coronavirus, SARS-CoV-2
James Zhu, Jiwoong Kim, Xue Xiao, Yunguan Wang, Danni Luo, Shuang Jiang, Ran Chen, Lin Xu, He Zhang, Lenny Moise, … Yang Xie Cold Spring Harbor Laboratory (2020-09-04) https://doi.org/ggq628
DOI: 10.1101/2020.02.08.939553 · PMID: 32908981
1259. Clinical Course and Outcomes of Critically Ill Patients With Middle East Respiratory Syndrome Coronavirus Infection
Yaseen M. Arabi, Ahmed A. Arifi, Hanan H. Balkhy, Hani Najm, Abdulaziz S. Aldawood, Alaa Ghabashi, Hassan Hawa, Adel Alothman, Abdulaziz Khaldi, Basel Al Raiy Annals of Internal Medicine (2014-03-18) https://doi.org/ggptxw
DOI: 10.7326/m13-2486 · PMID: 24474051
1260. Neutrophil-to-Lymphocyte Ratio Predicts Severe Illness Patients with 2019 Novel Coronavirus in the Early Stage
Jingyuan Liu, Yao Liu, Pan Xiang, Lin Pu, Haofeng Xiong, Chuansheng Li, Ming Zhang, Jianbo Tan, Yanli Xu, Rui Song, … Xianbo Wang Cold Spring Harbor Laboratory (2020-02-12) https://doi.org/ggrbdx
DOI: 10.1101/2020.02.10.20021584
1261. Dysregulation of Immune Response in Patients With Coronavirus 2019 (COVID-19) in Wuhan, China
Chuan Qin, Luoqi Zhou, Ziwei Hu, Shuoqi Zhang, Sheng Yang, Yu Tao, Cuihong Xie, Ke Ma, Ke Shang, Wei Wang, Dai-Shi Tian Clinical Infectious Diseases (2020-08-01) https://doi.org/ggpxcf
DOI: 10.1093/cid/ciaa248 · PMID: 32161940 · PMCID: PMC7108125
1262. Characteristics of lymphocyte subsets and cytokines in peripheral blood of 123 hospitalized patients with 2019 novel coronavirus pneumonia (NCP)
Suxin Wan, Qingjie Yi, Shibing Fan, Jinglong Lv, Xianxiang Zhang, Lian Guo, Chunhui Lang, Qing Xiao, Kaihu Xiao, Zhengjun Yi, … Yongping Chen Cold Spring Harbor Laboratory (2020-02-12) https://doi.org/ggq63b
DOI: 10.1101/2020.02.10.20021832
1263. Longitudinal Characteristics of Lymphocyte Responses and Cytokine Profiles in the Peripheral Blood of SARS-CoV-2 Infected Patients
Jing Liu, Sumeng Li, Jia Liu, Boyun Liang, Xiaobei Wang, Wei Li, Hua Wang, Qiaoxia Tong, Jianhua Yi, Lei Zhao, … Xin Zheng SSRN Electronic Journal (2020) https://doi.org/ggq655
DOI: 10.2139/ssrn.3539682
1264. Epidemiological and Clinical Characteristics of 17 Hospitalized Patients with 2019 Novel Coronavirus Infections Outside Wuhan, China
Jie Li, Shilin Li, Yurui Cai, Qin Liu, Xue Li, Zhaoping Zeng, Yanpeng Chu, Fangcheng Zhu, Fanxin Zeng Cold Spring Harbor Laboratory (2020-02-12) https://doi.org/ggq63c
DOI: 10.1101/2020.02.11.20022053
1265. ACE2 Expression in Kidney and Testis May Cause Kidney and Testis Damage After 2019-nCoV Infection
Caibin Fan, Kai Li, Yanhong Ding, Wei Lu, Jianqing Wang Cold Spring Harbor Laboratory (2020-02-13) https://doi.org/ggq63d
DOI: 10.1101/2020.02.12.20022418
1266. Aberrant pathogenic GM-CSF + T cells and inflammatory CD14 + CD16 + monocytes in severe pulmonary syndrome patients of a new coronavirus
Yonggang Zhou, Binqing Fu, Xiaohu Zheng, Dongsheng Wang, Changcheng Zhao, Yingjie qi, Rui Sun, Zhigang Tian, Xiaoling Xu, Haiming Wei Cold Spring Harbor Laboratory (2020-02-20) https://doi.org/ggq63f
DOI: 10.1101/2020.02.12.945576
1267. Clinical Characteristics of 2019 Novel Infected Coronavirus Pneumonia: A Systemic Review and Meta-analysis
Kai Qian, Yi Deng, Yong-Hang Tai, Jun Peng, Hao Peng, Li-Hong Jiang Cold Spring Harbor Laboratory (2020-02-17) https://doi.org/ggrgbq
DOI: 10.1101/2020.02.14.20021535
1268. Longitudinal characteristics of lymphocyte responses and cytokine profiles in the peripheral blood of SARS-CoV-2 infected patients
Jing Liu, Sumeng Li, Jia Liu, Boyun Liang, Xiaobei Wang, Hua Wang, Wei Li, Qiaoxia Tong, Jianhua Yi, Lei Zhao, … Xin Zheng Cold Spring Harbor Laboratory (2020-02-22) https://doi.org/ggq63g
DOI: 10.1101/2020.02.16.20023671
1269. Clinical and immunologic features in severe and moderate forms of Coronavirus Disease 2019
Guang Chen, Di Wu, Wei Guo, Yong Cao, Da Huang, Hongwu Wang, Tao Wang, Xiaoyun Zhang, Huilong Chen, Haijing Yu, … Qin Ning Cold Spring Harbor Laboratory (2020-02-19) https://doi.org/ggq63h
DOI: 10.1101/2020.02.16.20023903
1270. Protection of Rhesus Macaque from SARS-Coronavirus challenge by recombinant adenovirus vaccine
Yiyou Chen, Qiang Wei, Ruobing Li, Hong Gao, Hua Zhu, Wei Deng, Linlin Bao, Wei Tong, Zhe Cong, Hong Jiang, Chuan Qin Cold Spring Harbor Laboratory (2020-02-21) https://doi.org/ggq63k
DOI: 10.1101/2020.02.17.951939
1271. Reduction and Functional Exhaustion of T Cells in Patients with Coronavirus Disease 2019 (COVID-19)
Bo Diao, Chenhui Wang, Yingjun Tan, Xiewan Chen, Ying Liu, Lifen Ning, Li Chen, Min Li, Yueping Liu, Gang Wang, … Yongwen Chen Cold Spring Harbor Laboratory (2020-02-20) https://doi.org/ggq63m
DOI: 10.1101/2020.02.18.20024364
1272. Clinical characteristics of 25 death cases with COVID-19: a retrospective review of medical records in a single medical center, Wuhan, China
Xun Li, Luwen Wang, Shaonan Yan, Fan Yang, Longkui Xiang, Jiling Zhu, Bo Shen, Zuojiong Gong Cold Spring Harbor Laboratory (2020-02-25) https://doi.org/ggq63n
DOI: 10.1101/2020.02.19.20025239
1273. SARS-CoV-2 infection does not significantly cause acute renal injury: an analysis of 116 hospitalized patients with COVID-19 in a single hospital, Wuhan, China
Luwen Wang, Xun Li, Hui Chen, Shaonan Yan, Yan Li, Dong Li, Zuojiong Gong Cold Spring Harbor Laboratory (2020-02-23) https://doi.org/ggq63p
DOI: 10.1101/2020.02.19.20025288
1274. Clinical Characteristics of 138 Hospitalized Patients With 2019 Novel Coronavirus–Infected Pneumonia in Wuhan, China
Dawei Wang, Bo Hu, Chang Hu, Fangfang Zhu, Xing Liu, Jing Zhang, Binbin Wang, Hui Xiang, Zhenshun Cheng, Yong Xiong, … Zhiyong Peng JAMA (2020-03-17) https://doi.org/ggkh48
DOI: 10.1001/jama.2020.1585 · PMID: 32031570 · PMCID: PMC7042881
1275. Clinical characteristics of 2019 novel coronavirus infection in China
Wei-jie Guan, Zheng-yi Ni, Yu Hu, Wen-hua Liang, Chun-quan Ou, Jian-xing He, Lei Liu, Hong Shan, Chun-liang Lei, David S. C. Hui, … Nan-shan Zhong Cold Spring Harbor Laboratory (2020-02-09) https://doi.org/ggkj9s
DOI: 10.1101/2020.02.06.20020974
1276. Potential T-cell and B-cell Epitopes of 2019-nCoV
Ethan Fast, Russ B. Altman, Binbin Chen Cold Spring Harbor Laboratory (2020-03-18) https://doi.org/ggq63q
DOI: 10.1101/2020.02.19.955484
1277. Structure, function and antigenicity of the SARS-CoV-2 spike glycoprotein
Alexandra C. Walls, Young-Jun Park, M. Alexandra Tortorici, Abigail Wall, Andrew T. McGuire, David Veesler Cold Spring Harbor Laboratory (2020-02-20) https://doi.org/ggrgbr
DOI: 10.1101/2020.02.19.956581
1278. Breadth of concomitant immune responses underpinning viral clearance and patient recovery in a non-severe case of COVID-19
Irani Thevarajan, Thi HO Nguyen, Marios Koutsakos, Julian Druce, Leon Caly, Carolien E van de Sandt, Xiaoxiao Jia, Suellen Nicholson, Mike Catton, Benjamin Cowie, … Katherine Kedzierska Cold Spring Harbor Laboratory (2020-02-23) https://doi.org/ggq63r
DOI: 10.1101/2020.02.20.20025841
1279. The landscape of lung bronchoalveolar immune cells in COVID-19 revealed by single-cell RNA sequencing
Minfeng Liao, Yang Liu, Jin Yuan, Yanling Wen, Gang Xu, Juanjuan Zhao, Lin Chen, Jinxiu Li, Xin Wang, Fuxiang Wang, … Zheng Zhang Cold Spring Harbor Laboratory (2020-02-26) https://doi.org/ggq63s
DOI: 10.1101/2020.02.23.20026690
1280. Influenza A Virus Infection Induces Hyperresponsiveness in Human Lung Tissue-Resident and Peripheral Blood NK Cells
Marlena Scharenberg, Sindhu Vangeti, Eliisa Kekäläinen, Per Bergman, Mamdoh Al-Ameri, Niclas Johansson, Klara Sondén, Sara Falck-Jones, Anna Färnert, Hans-Gustaf Ljunggren, … Nicole Marquardt Frontiers in Immunology (2019-05-17) https://doi.org/ggq656
DOI: 10.3389/fimmu.2019.01116 · PMID: 31156653 · PMCID: PMC6534051
1281. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China
Chaolin Huang, Yeming Wang, Xingwang Li, Lili Ren, Jianping Zhao, Yi Hu, Li Zhang, Guohui Fan, Jiuyang Xu, Xiaoying Gu, … Bin Cao The Lancet (2020-02) https://doi.org/ggjfnn
DOI: 10.1016/s0140-6736(20)30183-5
1282. Alveolar Macrophages in the Resolution of Inflammation, Tissue Repair, and Tolerance to Infection
Benoit Allard, Alice Panariti, James G. Martin Frontiers in Immunology (2018-07-31) https://doi.org/gd3bnz
DOI: 10.3389/fimmu.2018.01777 · PMID: 30108592 · PMCID: PMC6079255
1283. PPAR-γ in Macrophages Limits Pulmonary Inflammation and Promotes Host Recovery following Respiratory Viral Infection
Su Huang, Bibo Zhu, In Su Cheon, Nick P. Goplen, Li Jiang, Ruixuan Zhang, R. Stokes Peebles, Matthias Mack, Mark H. Kaplan, Andrew H. Limper, Jie Sun Journal of Virology (2019-04-17) https://doi.org/ggq652
DOI: 10.1128/jvi.00030-19 · PMID: 30787149 · PMCID: PMC6475778
1284. Can routine laboratory tests discriminate 2019 novel coronavirus infected pneumonia from other community-acquired pneumonia?
Yunbao Pan, Guangming Ye, Xiantao Zeng, Guohong Liu, Xiaojiao Zeng, Xianghu Jiang, Jin Zhao, Liangjun Chen, Shuang Guo, Qiaoling Deng, … Xinghuan Wang Cold Spring Harbor Laboratory (2020-02-25) https://doi.org/ggq63t
DOI: 10.1101/2020.02.25.20024711
1285. Correlation Analysis Between Disease Severity and Inflammation-related Parameters in Patients with COVID-19 Pneumonia
Jing Gong, Hui Dong, Qingsong Xia, Zhaoyi Huang, Dingkun Wang, Yan Zhao, Wenhua Liu, Shenghao Tu, Mingmin Zhang, Qi Wang, Fuer Lu Cold Spring Harbor Laboratory (2020-02-26) https://doi.org/ggq63v
DOI: 10.1101/2020.02.25.20025643
1286. An Effective CTL Peptide Vaccine for Ebola Zaire Based on Survivors’ CD8+ Targeting of a Particular Nucleocapsid Protein Epitope with Potential Implications for COVID-19 Vaccine Design
CV Herst, S Burkholz, J Sidney, A Sette, PE Harris, S Massey, T Brasel, E Cunha-Neto, DS Rosa, WCH Chao, … R Rubsamen Cold Spring Harbor Laboratory (2020-04-06) https://doi.org/ggq63x
DOI: 10.1101/2020.02.25.963546
1287. Epitope-based peptide vaccine design and target site characterization against novel coronavirus disease caused by SARS-CoV-2
Lin Li, Ting Sun, Yufei He, Wendong Li, Yubo Fan, Jing Zhang Cold Spring Harbor Laboratory (2020-02-27) https://doi.org/ggnqwt
DOI: 10.1101/2020.02.25.965434
1288. The definition and risks of Cytokine Release Syndrome-Like in 11 COVID-19-Infected Pneumonia critically ill patients: Disease Characteristics and Retrospective Analysis
Wang Wenjun, Liu Xiaoqing, Wu Sipei, Lie Puyi, Huang Liyan, Li Yimin, Cheng Linling, Chen Sibei, Nong Lingbo, Lin Yongping, He Jianxing Cold Spring Harbor Laboratory (2020-02-27) https://doi.org/ggrgbs
DOI: 10.1101/2020.02.26.20026989
1289. Clinical characteristics of 36 non-survivors with COVID-19 in Wuhan, China
Ying Huang, Rui Yang, Ying Xu, Ping Gong Cold Spring Harbor Laboratory (2020-03-05) https://doi.org/ggq63z
DOI: 10.1101/2020.02.27.20029009
1290. Risk factors related to hepatic injury in patients with corona virus disease 2019
Lu Li, Shuang Li, Manman Xu, Pengfei Yu, Sujun Zheng, Zhongping Duan, Jing Liu, Yu Chen, Junfeng Li Cold Spring Harbor Laboratory (2020-03-10) https://doi.org/ggq632
DOI: 10.1101/2020.02.28.20028514
1291. Detectable serum SARS-CoV-2 viral load (RNAaemia) is closely associated with drastically elevated interleukin 6 (IL-6) level in critically ill COVID-19 patients
Xiaohua Chen, Binghong Zhao, Yueming Qu, Yurou Chen, Jie Xiong, Yong Feng, Dong Men, Qianchuan Huang, Ying Liu, Bo Yang, … Feng Li Cold Spring Harbor Laboratory (2020-03-03) https://doi.org/ggq633
DOI: 10.1101/2020.02.29.20029520
1293. Lymphopenia predicts disease severity of COVID-19: a descriptive and predictive study
Li Tan, Qi Wang, Duanyang Zhang, Jinya Ding, Qianchuan Huang, Yi-Quan Tang, Qiongshu Wang, Hongming Miao Cold Spring Harbor Laboratory (2020-03-03) https://doi.org/ggq634
DOI: 10.1101/2020.03.01.20029074
1294. The potential role of IL-6 in monitoring severe case of coronavirus disease 2019
Tao Liu, Jieying Zhang, Yuhui Yang, Hong Ma, Zhengyu Li, Jiaoyu Zhang, Ji Cheng, Xiaoyun Zhang, Yanxia Zhao, Zihan Xia, … Jianhua Yi Cold Spring Harbor Laboratory (2020-03-10) https://doi.org/ggq635
DOI: 10.1101/2020.03.01.20029769
1295. Clinical and Laboratory Profiles of 75 Hospitalized Patients with Novel Coronavirus Disease 2019 in Hefei, China
Zonghao Zhao, Jiajia Xie, Ming Yin, Yun Yang, Hongliang He, Tengchuan Jin, Wenting Li, Xiaowu Zhu, Jing Xu, Changcheng Zhao, … Xiaoling Ma Cold Spring Harbor Laboratory (2020-03-06) https://doi.org/ggq636
DOI: 10.1101/2020.03.01.20029785
1296. Exuberant elevation of IP-10, MCP-3 and IL-1ra during SARS-CoV-2 infection is associated with disease severity and fatal outcome
Yang Yang, Chenguang Shen, Jinxiu Li, Jing Yuan, Minghui Yang, Fuxiang Wang, Guobao Li, Yanjie Li, Li Xing, Ling Peng, … Yingxia Liu Cold Spring Harbor Laboratory (2020-03-06) https://doi.org/ggq637
DOI: 10.1101/2020.03.02.20029975
1297. Antibody responses to SARS-CoV-2 in patients of novel coronavirus disease 2019
Juanjuan Zhao, Quan Yuan, Haiyan Wang, Wei Liu, Xuejiao Liao, Yingying Su, Xin Wang, Jing Yuan, Tingdong Li, Jinxiu Li, … Zheng Zhang Cold Spring Harbor Laboratory (2020-03-02) https://doi.org/ggrbj6
DOI: 10.1101/2020.03.02.20030189
1298. Restoration of leukomonocyte counts is associated with viral clearance in COVID-19 hospitalized patients
Xiaoping Chen, Jiaxin Ling, Pingzheng Mo, Yongxi Zhang, Qunqun Jiang, Zhiyong Ma, Qian Cao, Wenjia Hu, Shi Zou, Liangjun Chen, … Yong Xiong Cold Spring Harbor Laboratory (2020-03-06) https://doi.org/ggq639
DOI: 10.1101/2020.03.03.20030437
1299. Effects of Systemically Administered Hydrocortisone on the Human Immunome
Matthew J. Olnes, Yuri Kotliarov, Angélique Biancotto, Foo Cheung, Jinguo Chen, Rongye Shi, Huizhi Zhou, Ena Wang, John S. Tsang, Robert Nussenblatt, The CHI Consortium Scientific Reports (2016-03-14) https://doi.org/f8dmvw
DOI: 10.1038/srep23002 · PMID: 26972611 · PMCID: PMC4789739
1301. Clinical findings in critically ill patients infected with SARS-CoV-2 in Guangdong Province, China: a multi-center, retrospective, observational study
Yonghao Xu, Zhiheng Xu, Xuesong Liu, Lihua Cai, Haichong Zheng, Yongbo Huang, Lixin Zhou, Linxi Huang, Yun Lin, Liehua Deng, … Yimin Li Cold Spring Harbor Laboratory (2020-03-06) https://doi.org/ggq64b
DOI: 10.1101/2020.03.03.20030668
1302. Multi-epitope vaccine design using an immunoinformatics approach for 2019 novel coronavirus (SARS-CoV-2)
Ye Feng, Min Qiu, Liang Liu, Shengmei Zou, Yun Li, Kai Luo, Qianpeng Guo, Ning Han, Yingqiang Sun, Kui Wang, … Fan Mo Cold Spring Harbor Laboratory (2020-06-30) https://doi.org/ggq64c
DOI: 10.1101/2020.03.03.962332
1303. Clinical Features of Patients Infected with the 2019 Novel Coronavirus (COVID-19) in Shanghai, China
Min Cao, Dandan Zhang, Youhua Wang, Yunfei Lu, Xiangdong Zhu, Ying Li, Honghao Xue, Yunxiao Lin, Min Zhang, Yiguo Sun, … Hongzhou Lu Cold Spring Harbor Laboratory (2020-03-06) https://doi.org/ggq64d
DOI: 10.1101/2020.03.04.20030395 · PMID: 32511465
1304. Serological detection of 2019-nCoV respond to the epidemic: A useful complement to nucleic acid testing
Jin Zhang, Jianhua Liu, Na Li, Yong Liu, Rui Ye, Xiaosong Qin, Rui Zheng Cold Spring Harbor Laboratory (2020-03-06) https://doi.org/ggq64f
DOI: 10.1101/2020.03.04.20030916
1305. Human Kidney is a Target for Novel Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Infection
Bo Diao, Chenhui Wang, Rongshuai Wang, Zeqing Feng, Yingjun Tan, Huiming Wang, Changsong Wang, Liang Liu, Ying Liu, Yueping Liu, … Yongwen Chen Cold Spring Harbor Laboratory (2020-04-10) https://doi.org/ggq64g
DOI: 10.1101/2020.03.04.20031120
1306. COVID-19 early warning score: a multi-parameter screening tool to identify highly suspected patients
Cong-Ying Song, Jia Xu, Jian-Qin He, Yuan-Qiang Lu Cold Spring Harbor Laboratory (2020-03-08) https://doi.org/ggq64h
DOI: 10.1101/2020.03.05.20031906
1307. LY6E impairs coronavirus fusion and confers immune control of viral disease
Stephanie Pfaender, Katrina B. Mar, Eleftherios Michailidis, Annika Kratzel, Dagny Hirt, Philip V’kovski, Wenchun Fan, Nadine Ebert, Hanspeter Stalder, Hannah Kleine-Weber, … Volker Thiel Cold Spring Harbor Laboratory (2020-03-07) https://doi.org/dpvn
DOI: 10.1101/2020.03.05.979260 · PMID: 32511345
1308. A preliminary study on serological assay for severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) in 238 admitted hospital patients
Lei Liu, Wanbing Liu, Yaqiong Zheng, Xiaojing Jiang, Guomei Kou, Jinya Ding, Qiongshu Wang, Qianchuan Huang, Yinjuan Ding, Wenxu Ni, … Shangen Zheng Cold Spring Harbor Laboratory (2020-03-08) https://doi.org/ggq64j
DOI: 10.1101/2020.03.06.20031856
1309. Monoclonal antibodies for the S2 subunit of spike of SARS-CoV cross-react with the newly-emerged SARS-CoV-2
Zhiqiang Zheng, Vanessa M. Monteil, Sebastian Maurer-Stroh, Chow Wenn Yew, Carol Leong, Nur Khairiah Mohd-Ismail, Suganya Cheyyatraivendran Arularasu, Vincent Tak Kwong Chow, Raymond Lin Tzer Pin, Ali Mirazimi, … Yee-Joo Tan Cold Spring Harbor Laboratory (2020-03-07) https://doi.org/ggrbj7
DOI: 10.1101/2020.03.06.980037
1310. Mortality of COVID-19 is Associated with Cellular Immune Function Compared to Immune Function in Chinese Han Population
Qiang Zeng, Yong-zhe Li, Gang Huang, Wei Wu, Sheng-yong Dong, Yang Xu Cold Spring Harbor Laboratory (2020-03-13) https://doi.org/ggq64k
DOI: 10.1101/2020.03.08.20031229
1311. Retrospective Analysis of Clinical Features in 101 Death Cases with COVID-19
Hua Fan, Lin Zhang, Bin Huang, Muxin Zhu, Yong Zhou, Huan Zhang, Xiaogen Tao, Shaohui Cheng, Wenhu Yu, Liping Zhu, Jian Chen Cold Spring Harbor Laboratory (2020-03-17) https://doi.org/ggq64n
DOI: 10.1101/2020.03.09.20033068
1312. Relationship between the ABO Blood Group and the COVID-19 Susceptibility
Jiao Zhao, Yan Yang, Hanping Huang, Dong Li, Dongfeng Gu, Xiangfeng Lu, Zheng Zhang, Lei Liu, Ting Liu, Yukun Liu, … Peng George Wang Cold Spring Harbor Laboratory (2020-03-27) https://doi.org/ggpn3d
DOI: 10.1101/2020.03.11.20031096
1314. Immune phenotyping based on neutrophil-to-lymphocyte ratio and IgG predicts disease severity and outcome for patients with COVID-19
Bicheng Zhang, Xiaoyang Zhou, Chengliang Zhu, Fan Feng, Yanru Qiu, Jia Feng, Qingzhu Jia, Qibin Song, Bo Zhu, Jun Wang Cold Spring Harbor Laboratory (2020-03-16) https://doi.org/ggq64q
DOI: 10.1101/2020.03.12.20035048
1315. Lack of Reinfection in Rhesus Macaques Infected with SARS-CoV-2
Linlin Bao, Wei Deng, Hong Gao, Chong Xiao, Jiayi Liu, Jing Xue, Qi Lv, Jiangning Liu, Pin Yu, Yanfeng Xu, … Chuan Qin Cold Spring Harbor Laboratory (2020-05-01) https://doi.org/ggn8r8
DOI: 10.1101/2020.03.13.990226
1316. A highly conserved cryptic epitope in the receptor-binding domains of SARS-CoV-2 and SARS-CoV
Meng Yuan, Nicholas C. Wu, Xueyong Zhu, Chang-Chun D. Lee, Ray T. Y. So, Huibin Lv, Chris K. P. Mok, Ian A. Wilson Cold Spring Harbor Laboratory (2020-03-14) https://doi.org/ggq64s
DOI: 10.1101/2020.03.13.991570
1317. Highly accurate and sensitive diagnostic detection of SARS-CoV-2 by digital PCR
Lianhua Dong, Junbo Zhou, Chunyan Niu, Quanyi Wang, Yang Pan, Sitong Sheng, Xia Wang, Yongzhuo Zhang, Jiayi Yang, Manqing Liu, … Xiang Fang Cold Spring Harbor Laboratory (2020-03-30) https://doi.org/ggqnqh
DOI: 10.1101/2020.03.14.20036129
1318. SARS-CoV-2 invades host cells via a novel route: CD147-spike protein
Ke Wang, Wei Chen, Yu-Sen Zhou, Jian-Qi Lian, Zheng Zhang, Peng Du, Li Gong, Yang Zhang, Hong-Yong Cui, Jie-Jie Geng, … Zhi-Nan Chen Cold Spring Harbor Laboratory (2020-03-14) https://doi.org/ggq64t
DOI: 10.1101/2020.03.14.988345
1319. CD147 (EMMPRIN/Basigin) in kidney diseases: from an inflammation and immune system viewpoint
Tomoki Kosugi, Kayaho Maeda, Waichi Sato, Shoichi Maruyama, Kenji Kadomatsu Nephrology Dialysis Transplantation (2015-07) https://doi.org/ggq624
DOI: 10.1093/ndt/gfu302 · PMID: 25248362
1321. Cancer-related issues of CD147.
Ulrich H Weidle, Werner Scheuer, Daniela Eggle, Stefan Klostermann, Hannes Stockinger Cancer genomics & proteomicshttps://www.ncbi.nlm.nih.gov/pubmed/20551248
PMID: 20551248
1322. Blood single cell immune profiling reveals the interferon-MAPK pathway mediated adaptive immune response for COVID-19
Lulin Huang, Yi Shi, Bo Gong, Li Jiang, Xiaoqi Liu, Jialiang Yang, Juan Tang, Chunfang You, Qi Jiang, Bo Long, … Zhenglin Yang Cold Spring Harbor Laboratory (2020-03-17) https://doi.org/ggq64v
DOI: 10.1101/2020.03.15.20033472
1323. Cross-reactive antibody response between SARS-CoV-2 and SARS-CoV infections
Huibin Lv, Nicholas C. Wu, Owen Tak-Yin Tsang, Meng Yuan, Ranawaka A. P. M. Perera, Wai Shing Leung, Ray T. Y. So, Jacky Man Chun Chan, Garrick K. Yip, Thomas Shiu Hong Chik, … Chris K. P. Mok Cold Spring Harbor Laboratory (2020-03-17) https://doi.org/ggq64w
DOI: 10.1101/2020.03.15.993097 · PMID: 32511317
1324. The feasibility of convalescent plasma therapy in severe COVID- 19 patients: a pilot study
Kai Duan, Bende Liu, Cesheng Li, Huajun Zhang, Ting Yu, Jieming Qu, Min Zhou, Li Chen, Shengli Meng, Yong Hu, … Xiaoming Yang Cold Spring Harbor Laboratory (2020-03-23) https://doi.org/dqrs
DOI: 10.1101/2020.03.16.20036145
1325. Hydroxychloroquine and azithromycin as a treatment of COVID-19: results of an open-label non-randomized clinical trial
Philippe Gautret, Jean-Christophe Lagier, Philippe Parola, Van Thuan Hoang, Line Meddeb, Morgane Mailhe, Barbara Doudier, Johan Courjon, Valérie Giordanengo, Vera Esteves Vieira, … Didier Raoult Cold Spring Harbor Laboratory (2020-03-20) https://doi.org/dqbv
DOI: 10.1101/2020.03.16.20037135
1326. Chloroquine: Modes of action of an undervalued drug
Rodolfo Thomé, Stefanie Costa Pinto Lopes, Fabio Trindade Maranhão Costa, Liana Verinaud Immunology Letters (2013-06) https://doi.org/f5b5cr
DOI: 10.1016/j.imlet.2013.07.004 · PMID: 23891850
1327. Patients with LRBA deficiency show CTLA4 loss and immune dysregulation responsive to abatacept therapy
B. Lo, K. Zhang, W. Lu, L. Zheng, Q. Zhang, C. Kanellopoulou, Y. Zhang, Z. Liu, J. M. Fritz, R. Marsh, … M. B. Jordan Science (2015-07-23) https://doi.org/f7kc8d
DOI: 10.1126/science.aaa1663 · PMID: 26206937
1328. The sequence of human ACE2 is suboptimal for binding the S spike protein of SARS coronavirus 2
Erik Procko Cold Spring Harbor Laboratory (2020-05-11) https://doi.org/ggrbj8
DOI: 10.1101/2020.03.16.994236 · PMID: 32511321
1329. Comparative Pathogenesis Of COVID-19, MERS And SARS In A Non-Human Primate Model
Barry Rockx, Thijs Kuiken, Sander Herfst, Theo Bestebroer, Mart M. Lamers, Dennis de Meulder, Geert van Amerongen, Judith van den Brand, Nisreen M. A. Okba, Debby Schipper, … Bart L. Haagmans Cold Spring Harbor Laboratory (2020-03-17) https://doi.org/ggq649
DOI: 10.1101/2020.03.17.995639
1330. Lethal Infection of K18-hACE2 Mice Infected with Severe Acute Respiratory Syndrome Coronavirus
Paul B. McCray, Lecia Pewe, Christine Wohlford-Lenane, Melissa Hickey, Lori Manzel, Lei Shi, Jason Netland, Hong Peng Jia, Carmen Halabi, Curt D. Sigmund, … Stanley Perlman Journal of Virology (2007-01-15) https://doi.org/b2dr3s
DOI: 10.1128/jvi.02012-06 · PMID: 17079315 · PMCID: PMC1797474
1331. Modeling the Impact of Asymptomatic Carriers on COVID-19 Transmission Dynamics During Lockdown
Jacob B. Aguilar, Jeremy Samuel Faust, Lauren M. Westafer, Juan B. Gutierrez Cold Spring Harbor Laboratory (2020-08-11) https://doi.org/ggqnvp
DOI: 10.1101/2020.03.18.20037994
1332. Antibody responses to SARS-CoV-2 in COVID-19 patients: the perspective application of serological tests in clinical practice
Quan-xin Long, Hai-jun Deng, Juan Chen, Jie-li Hu, Bei-zhong Liu, Pu Liao, Yong Lin, Li-hua Yu, Zhan Mo, Yin-yin Xu, … Ai-long Huang Cold Spring Harbor Laboratory (2020-03-20) https://doi.org/ggpvz3
DOI: 10.1101/2020.03.18.20038018
1333. Heat inactivation of serum interferes with the immunoanalysis of antibodies to SARS-CoV-2
Xiumei Hu, Taixue An, Bo Situ, Yuhai Hu, Zihao Ou, Qiang Li, Xiaojing He, Ye Zhang, Peifu Tian, Dehua Sun, … Lei Zheng Cold Spring Harbor Laboratory (2020-03-16) https://doi.org/ggq646
DOI: 10.1101/2020.03.12.20034231
1334. SARS-CoV-2 specific antibody responses in COVID-19 patients
Nisreen M. A. Okba, Marcel A. Müller, Wentao Li, Chunyan Wang, Corine H. GeurtsvanKessel, Victor M. Corman, Mart M. Lamers, Reina S. Sikkema, Erwin de Bruin, Felicity D. Chandler, … Bart L. Haagmans Cold Spring Harbor Laboratory (2020-03-20) https://doi.org/ggpvz2
DOI: 10.1101/2020.03.18.20038059
1335. A brief review of antiviral drugs evaluated in registered clinical trials for COVID-19
Drifa Belhadi, Nathan Peiffer-Smadja, François-Xavier Lescure, Yazdan Yazdanpanah, France Mentré, Cédric Laouénan Cold Spring Harbor Laboratory (2020-03-27) https://doi.org/ggq65b
DOI: 10.1101/2020.03.18.20038190
1336. ACE-2 Expression in the Small Airway Epithelia of Smokers and COPD Patients: Implications for COVID-19
Janice M. Leung, Chen X. Yang, Anthony Tam, Tawimas Shaipanich, Tillie-Louise Hackett, Gurpreet K. Singhera, Delbert R. Dorscheid, Don D. Sin Cold Spring Harbor Laboratory (2020-03-23) https://doi.org/dqx2
DOI: 10.1101/2020.03.18.20038455
1338. Association between Clinical, Laboratory and CT Characteristics and RT-PCR Results in the Follow-up of COVID-19 patients
Hang Fu, Huayan Xu, Na Zhang, Hong Xu, Zhenlin Li, Huizhu Chen, Rong Xu, Ran Sun, Lingyi Wen, Linjun Xie, … Yingkun Guo Cold Spring Harbor Laboratory (2020-03-23) https://doi.org/ggq65c
DOI: 10.1101/2020.03.19.20038315
1339. An orally bioavailable broad-spectrum antiviral inhibits SARS-CoV-2 and multiple endemic, epidemic and bat coronavirus
Timothy P. Sheahan, Amy C. Sims, Shuntai Zhou, Rachel L. Graham, Collin S. Hill, Sarah R. Leist, Alexandra Schäfer, Kenneth H. Dinnon, Stephanie A. Montgomery, Maria L. Agostini, … Ralph S. Baric Cold Spring Harbor Laboratory (2020-03-20) https://doi.org/ggrbkb
DOI: 10.1101/2020.03.19.997890
1340. Identification of antiviral drug candidates against SARS-CoV-2 from FDA-approved drugs
Sangeun Jeon, Meehyun Ko, Jihye Lee, Inhee Choi, Soo Young Byun, Soonju Park, David Shum, Seungtaek Kim Cold Spring Harbor Laboratory (2020-03-28) https://doi.org/ggq65h
DOI: 10.1101/2020.03.20.999730
1341. Respiratory disease and virus shedding in rhesus macaques inoculated with SARS-CoV-2
Vincent J. Munster, Friederike Feldmann, Brandi N. Williamson, Neeltje van Doremalen, Lizzette Pérez-Pérez, Jonathan Schulz, Kimberly Meade-White, Atsushi Okumura, Julie Callison, Beniah Brumbaugh, … Emmie de Wit Cold Spring Harbor Laboratory (2020-03-21) https://doi.org/ggq65j
DOI: 10.1101/2020.03.21.001628 · PMID: 32511299
1342. Ocular conjunctival inoculation of SARS-CoV-2 can cause mild COVID-19 in Rhesus macaques
Wei Deng, Linlin Bao, Hong Gao, Zhiguang Xiang, Yajin Qu, Zhiqi Song, Shunran Gong, Jiayi Liu, Jiangning Liu, Pin Yu, … Chuan Qin Cold Spring Harbor Laboratory (2020-03-30) https://doi.org/ggq64r
DOI: 10.1101/2020.03.13.990036
1343. ACE2 Expression is Increased in the Lungs of Patients with Comorbidities Associated with Severe COVID-19
Bruna G. G. Pinto, Antonio E. R. Oliveira, Youvika Singh, Leandro Jimenez, Andre N A. Gonçalves, Rodrigo L. T. Ogava, Rachel Creighton, Jean Pierre Schatzmann Peron, Helder I. Nakaya Cold Spring Harbor Laboratory (2020-03-27) https://doi.org/ggq65k
DOI: 10.1101/2020.03.21.20040261 · PMID: 32511627
1345. CD147 facilitates HIV-1 infection by interacting with virus-associated cyclophilin A
T. Pushkarsky, G. Zybarth, L. Dubrovsky, V. Yurchenko, H. Tang, H. Guo, B. Toole, B. Sherry, M. Bukrinsky Proceedings of the National Academy of Sciences (2001-05-15) https://doi.org/cc4c7p
DOI: 10.1073/pnas.111583198 · PMID: 11353871 · PMCID: PMC33473
1346. CD147/EMMPRIN Acts as a Functional Entry Receptor for Measles Virus on Epithelial Cells
Akira Watanabe, Misako Yoneda, Fusako Ikeda, Yuri Terao-Muto, Hiroki Sato, Chieko Kai Journal of Virology (2010-05-01) https://doi.org/dpcsqg
DOI: 10.1128/jvi.02168-09 · PMID: 20147391 · PMCID: PMC2863760
1347. Basigin is a receptor essential for erythrocyte invasion by Plasmodium falciparum
Cécile Crosnier, Leyla Y. Bustamante, S. Josefin Bartholdson, Amy K. Bei, Michel Theron, Makoto Uchikawa, Souleymane Mboup, Omar Ndir, Dominic P. Kwiatkowski, Manoj T. Duraisingh, … Gavin J. Wright Nature (2011-11-09) https://doi.org/dm59hf
DOI: 10.1038/nature10606 · PMID: 22080952 · PMCID: PMC3245779
1348. Function of HAb18G/CD147 in Invasion of Host Cells by Severe Acute Respiratory Syndrome Coronavirus
Zhinan Chen, Li Mi, Jing Xu, Jiyun Yu, Xianhui Wang, Jianli Jiang, Jinliang Xing, Peng Shang, Airong Qian, Yu Li, … Ping Zhu The Journal of Infectious Diseases (2005-03) https://doi.org/cd8snd
DOI: 10.1086/427811 · PMID: 15688292 · PMCID: PMC7110046
1349. CD147 mediates intrahepatic leukocyte aggregation and determines the extent of liver injury
Christine Yee, Nathan M. Main, Alexandra Terry, Igor Stevanovski, Annette Maczurek, Alison J. Morgan, Sarah Calabro, Alison J. Potter, Tina L. Iemma, David G. Bowen, … Nicholas A. Shackel PLOS ONE (2019-07-10) https://doi.org/ggq654
DOI: 10.1371/journal.pone.0215557 · PMID: 31291257 · PMCID: PMC6619953
1350. Characterisation of the transcriptome and proteome of SARS-CoV-2 using direct RNA sequencing and tandem mass spectrometry reveals evidence for a cell passage induced in-frame deletion in the spike glycoprotein that removes the furin-like cleavage site
Andrew D. Davidson, Maia Kavanagh Williamson, Sebastian Lewis, Deborah Shoemark, Miles W. Carroll, Kate Heesom, Maria Zambon, Joanna Ellis, Phillip A. Lewis, Julian A. Hiscox, David A. Matthews Cold Spring Harbor Laboratory (2020-03-24) https://doi.org/ggq65n
DOI: 10.1101/2020.03.22.002204
1351. Modifications to the Hemagglutinin Cleavage Site Control the Virulence of a Neurotropic H1N1 Influenza Virus
Xiangjie Sun, Longping V. Tse, A Damon Ferguson, Gary R. Whittaker Journal of Virology (2010-09-01) https://doi.org/drs2zt
DOI: 10.1128/jvi.00797-10 · PMID: 20554779 · PMCID: PMC2919019
1352. The architecture of SARS-CoV-2 transcriptome
Dongwan Kim, Joo-Yeon Lee, Jeong-Sun Yang, Jun Won Kim, V. Narry Kim, Hyeshik Chang Cold Spring Harbor Laboratory (2020-03-14) https://doi.org/ggpx9q
DOI: 10.1101/2020.03.12.988865
1353. First Clinical Study Using HCV Protease Inhibitor Danoprevir to Treat Naïve and Experienced COVID-19 Patients
Hongyi Chen, Zhicheng Zhang, Li Wang, Zhihua Huang, Fanghua Gong, Xiaodong Li, Yahong Chen, Jinzi J. Wu Cold Spring Harbor Laboratory (2020-03-24) https://doi.org/ggrgbt
DOI: 10.1101/2020.03.22.20034041
1354. Preclinical Characteristics of the Hepatitis C Virus NS3/4A Protease Inhibitor ITMN-191 (R7227)
Scott D. Seiwert, Steven W. Andrews, Yutong Jiang, Vladimir Serebryany, Hua Tan, Karl Kossen, P. T. Ravi Rajagopalan, Shawn Misialek, Sarah K. Stevens, Antitsa Stoycheva, … Lawrence M. Blatt Antimicrobial Agents and Chemotherapy (2008-12) https://doi.org/btpg52
DOI: 10.1128/aac.00699-08 · PMID: 18824605 · PMCID: PMC2592891
1355. Efficacy and Safety of All-oral, 12-week Ravidasvir Plus Ritonavir-boosted Danoprevir and Ribavirin in Treatment-naïve Noncirrhotic HCV Genotype 1 Patients: Results from a Phase 2/3 Clinical Trial in China
Xiaoyuan Xu, Bo Feng, Yujuan Guan, Sujun Zheng, Jifang Sheng, Xingxiang Yang, Yuanji Ma, Yan Huang, Yi Kang, Xiaofeng Wen, … Lai Wei Journal of Clinical and Translational Hepatology (2019-09-30) https://doi.org/ggrbkd
DOI: 10.14218/jcth.2019.00033 · PMID: 31608212 · PMCID: PMC6783683
1357. Serology characteristics of SARS-CoV-2 infection since the exposure and post symptoms onset
Bin Lou, Ting-Dong Li, Shu-Fa Zheng, Ying-Ying Su, Zhi-Yong Li, Wei Liu, Fei Yu, Sheng-Xiang Ge, Qian-Da Zou, Quan Yuan, … Yu Chen Cold Spring Harbor Laboratory (2020-03-27) https://doi.org/ggrbkc
DOI: 10.1101/2020.03.23.20041707
1358. SARS-CoV-2 launches a unique transcriptional signature from in vitro, ex vivo, and in vivo systems
Daniel Blanco-Melo, Benjamin E. Nilsson-Payant, Wen-Chun Liu, Rasmus Møller, Maryline Panis, David Sachs, Randy A. Albrecht, Benjamin R. tenOever Cold Spring Harbor Laboratory (2020-03-24) https://doi.org/ggq65q
DOI: 10.1101/2020.03.24.004655
1359. A New Predictor of Disease Severity in Patients with COVID-19 in Wuhan, China
Ying Zhou, Zhen Yang, Yanan Guo, Shuang Geng, Shan Gao, Shenglan Ye, Yi Hu, Yafei Wang Cold Spring Harbor Laboratory (2020-03-27) https://doi.org/ggq65r
DOI: 10.1101/2020.03.24.20042119
1360. Metabolic disturbances and inflammatory dysfunction predict severity of coronavirus disease 2019 (COVID-19): a retrospective study
Shuke Nie, Xueqing Zhao, Kang Zhao, Zhaohui Zhang, Zhentao Zhang, Zhan Zhang Cold Spring Harbor Laboratory (2020-03-26) https://doi.org/ggq65s
DOI: 10.1101/2020.03.24.20042283
1361. Viral Kinetics and Antibody Responses in Patients with COVID-19
Wenting Tan, Yanqiu Lu, Juan Zhang, Jing Wang, Yunjie Dan, Zhaoxia Tan, Xiaoqing He, Chunfang Qian, Qiangzhong Sun, Qingli Hu, … Guohong Deng Cold Spring Harbor Laboratory (2020-03-26) https://doi.org/ggq65t
DOI: 10.1101/2020.03.24.20042382
1362. Global profiling of SARS-CoV-2 specific IgG/ IgM responses of convalescents using a proteome microarray
He-wei Jiang, Yang Li, Hai-nan Zhang, Wei Wang, Dong Men, Xiao Yang, Huan Qi, Jie Zhou, Sheng-ce Tao Cold Spring Harbor Laboratory (2020-03-27) https://doi.org/ggq65g
DOI: 10.1101/2020.03.20.20039495
1363. Anti–spike IgG causes severe acute lung injury by skewing macrophage responses during acute SARS-CoV infection
Li Liu, Qiang Wei, Qingqing Lin, Jun Fang, Haibo Wang, Hauyee Kwok, Hangying Tang, Kenji Nishiura, Jie Peng, Zhiwu Tan, … Zhiwei Chen JCI Insight (2019-02-21) https://doi.org/ggqbpw
DOI: 10.1172/jci.insight.123158 · PMID: 30830861 · PMCID: PMC6478436
1364. COVID-19 infection induces readily detectable morphological and inflammation-related phenotypic changes in peripheral blood monocytes, the severity of which correlate with patient outcome
Dan Zhang, Rui Guo, Lei Lei, Hongjuan Liu, Yawen Wang, Yili Wang, Hongbo Qian, Tongxin Dai, Tianxiao Zhang, Yanjun Lai, … Jinsong Hu Cold Spring Harbor Laboratory (2020-03-26) https://doi.org/ggq65v
DOI: 10.1101/2020.03.24.20042655
1365. Correlation between universal BCG vaccination policy and reduced morbidity and mortality for COVID-19: an epidemiological study
Aaron Miller, Mac Josh Reandelar, Kimberly Fasciglione, Violeta Roumenova, Yan Li, Gonzalo H. Otazu Cold Spring Harbor Laboratory (2020-03-28) https://doi.org/ggq65w
DOI: 10.1101/2020.03.24.20042937
1366. Non-specific effects of BCG vaccine on viral infections
S. J. C. F. M. Moorlag, R. J. W. Arts, R. van Crevel, M. G. Netea Clinical Microbiology and Infection (2019-12) https://doi.org/ggq62z
DOI: 10.1016/j.cmi.2019.04.020 · PMID: 31055165
1367. BCG vaccination to reduce the impact of COVID-19 in healthcare workers (The BRACE Trial)
Murdoch Children’s Research Institute https://www.mcri.edu.au/BRACE
1368. Non-neuronal expression of SARS-CoV-2 entry genes in the olfactory system suggests mechanisms underlying COVID-19-associated anosmia
David H. Brann, Tatsuya Tsukahara, Caleb Weinreb, Marcela Lipovsek, Koen Van den Berge, Boying Gong, Rebecca Chance, Iain C. Macaulay, Hsin-jung Chou, Russell Fletcher, … Sandeep Robert Datta Cold Spring Harbor Laboratory (2020-05-18) https://doi.org/ggqr4m
DOI: 10.1101/2020.03.25.009084
1369. Cigarette smoke exposure and inflammatory signaling increase the expression of the SARS-CoV-2 receptor ACE2 in the respiratory tract
Joan C. Smith, Erin L. Sausville, Vishruth Girish, Monet Lou Yuan, Kristen M. John, Jason M. Sheltzer Cold Spring Harbor Laboratory (2020-04-26) https://doi.org/ggq65x
DOI: 10.1101/2020.03.28.013672
1370. The comparative superiority of IgM-IgG antibody test to real-time reverse transcriptase PCR detection for SARS-CoV-2 infection diagnosis
Rui Liu, Xinghui Liu, Huan Han, Muhammad Adnan Shereen, Zhili Niu, Dong Li, Fang Liu, Kailang Wu, Zhen Luo, Chengliang Zhu Cold Spring Harbor Laboratory (2020-03-30) https://doi.org/ggqtp5
DOI: 10.1101/2020.03.28.20045765
13 Appendix A
This appendix contains reviews produced by the Immunology Institute of the Icahn School of Medicine
13.1 Potent binding of 2019 novel coronavirus spike protein by a SARS coronavirus-specific human monoclonal antibody
Considering the relatively high identity of the receptor binding domain
(RBD) of the spike proteins from 2019-nCoV and SARS-CoV (73%), this
study aims to assess the cross-reactivity of several anti-SARS-CoV
monoclonal antibodies with 2019-nCoV. The results showed that the
SARS-CoV-specific antibody CR3022 can potently bind 2019-nCoV RBD.
13.1.3 Main Findings
The structure of the 2019-nCoV spike RBD and its conformation in complex
with the receptor angiotensin-converting enzyme (ACE2) was modeled in
silico and compared with the SARS-CoV RBD structure. The models
predicted very similar RBD-ACE2 interactions for both viruses. The
binding capacity of representative SARS-CoV-RBD specific monoclonal
antibodies (m396, CR3014, and CR3022) to recombinant 2019-nCoV RBD was
then investigated by ELISA and their binding kinetics studied using
biolayer interferometry. The analysis showed that only CR3022 was able
to bind 2019-nCoV RBD with high affinity (KD of 6.3 nM), however it did
not interfere with ACE2 binding. Antibodies m396 and CR3014, which
target the ACE2 binding site of SARS-CoV failed to bind 2019-nCoV spike
protein.
13.1.4 Limitations
The 2019-nCoV RBD largely differ from the SARS-CoV at the C-terminus
residues, which drastically impact the cross-reactivity of antibodies
described for other B beta-coronaviruses, including SARS-CoV. This study
claims that CR3022 antibody could be a potential candidate for therapy.
However, none of the antibodies assayed in this work showed
cross-reactivity with the ACE2 binding site of 2019-nCoV, essential for
the replication of this virus. Furthermore, neutralization assays with
2019-nCoV virus or pseudovirus were not performed. Although the use of
neutralizing antibodies is an interesting approach, these results
suggest that it is critical the development of novel monoclonal
antibodies able to specifically bind 2019-nCoV spike protein.
13.1.5 Credit
Review by D.L.O as part of a project by students, postdocs and faculty
at the Immunology Institute of the Icahn School of Medicine, Mount
Sinai.
13.2 Integrative Bioinformatics Analysis Provides Insight into the Molecular Mechanisms of 2019-nCoV
The authors used bioinformatics tools to identify features
of ACE2 expression in the lungs of different patent groups: healthy,
smokers, patients with chronic airway disease (i.e., COPD) or asthma.
They used gene expression data publicly available from GEO that included
lung tissues, bronchoalveolar lavage, bronchial epithelial cells, small
airway epithelial cells, or SARS-Cov infected cells.
The authors describe no significant differences in ACE2 expression in
lung tissues of Healthy, COPD, and Asthma groups (p=0.85); or in BAL of
Healthy and COPD (p=0.48); or in epithelial brushings of Healthy and
Mild/Moderate/Severe Asthma (p=0.99). ACE2 was higher in the small
airway epithelium of long-term smokers vs non-smokers (p<0.001).
Consistently, there was a trend of higher ACE2 expression in the
bronchial airway epithelial cells 24h post-acute smoking exposure
(p=0.073). Increasing ACE2 expression at 24h and 48h compared to 12h
post SARS-Cov infection (p=0.026; n=3 at each time point) was also
detected.
15 lung samples’ data from healthy participants were separated into high
and low ACE2 expression groups. “High” ACE2 expression was associated
with the following GO pathways: innate and adaptive immune responses, B
cell mediated immunity, cytokine secretion, and IL-1, IL-10, IL-6, IL-8
cytokines. The authors speculate that a high basal ACE2 expression will
increase susceptibility to SARS-CoV infection.
In 3 samples SARS-Cov infection was associated with IL-1, IL-10 and IL-6
cytokine production (GO pathways) at 24h. And later, at 48h, with T-cell
activation and T-cell cytokine production. It is unclear whether those
changes were statistically significant.
The authors describe a time course quantification of immune infiltrates
in epithelial cells infected with SARS-Cov infection. They state that in
healthy donors ACE2 expression did not correlate with the immune cell
infiltration. However, in SARS-Cov samples, at 48h they found that ACE2
correlated with neutrophils, NK-, Th17-, Th2-, Th1- cells, and DCs.
Again, while authors claim significance, the corresponding correlation
coefficients and p-values are not presented in the text or figures. In
addition, the source of the data for this analysis is not clear.
Using network analysis, proteins SRC, FN1, MAPK3, LYN, MBP, NLRC4, NLRP1
and PRKCD were found to be central (Hub proteins) in the regulating
network of cytokine secretion after coronavirus infection. Authors
conclude this indicates that these molecules were critically important
in ACE2-induced inflammatory response. Additionally, authors speculate
that the increased expression of ACE2 affected RPS3 and SRC, which were
the two hub genes involved in viral replication and inflammatory
response.
13.2.3 Limitations
The methods section is very limited and does not
describe any of the statistical analyses; and description of the
construction of the regulatory protein networks is also limited. For the
findings in Figures 2 authors claim significance, which is not supported
by p-values or coefficients. For the sample selection, would be useful
if sample sizes and some of the patients’ demographics (e.g. age) were
described.
For the analysis of high vs low ACE2 expression in healthy subjects, it
is not clear what was the cut off for ‘high’ expression and how it was
determined. Additionally, further laboratory studies are warranted to
confirm that high ACE2 gene expression would have high correlation with
the amount of ACE2 protein on cell surface. For the GO pathway analysis
significance was set at p<0.05, but not adjusted for multiple
comparisons.
There were no samples with SARS-CoV-2 infection. While SARS-Cov and
SARS-CoV-2 both use ACE2 to enter the host cells, the analysis only
included data on SARS-Cov and any conclusions about SARS-CoV2 are
limited.
Upon checking GSE accession numbers of the datasets references, two
might not be cited correctly: GSE37758 (“A spergillus niger: Control
(fructose) vs. steam-exploded sugarcane induction (SEB)”" was used in
this paper as “lung tissue” data) and GSE14700 (“Steroid Pretreatment
of Organ Donors to Prevent Postischemic Renal Allograft Failure: A
Randomized, Controlled Trial” – was used as SARS-Cov infection data).
13.2.4 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.3 Diarrhea may be underestimated: a missing link in 2019 novel coronavirus
This study examined the incidence of diarrhea in
patients infected with SARS-CoV-2 across three recently published
cohorts and found that there are statistically significant differences
by Fisher’s exact test. They report that this could be due to subjective
diagnosis criterion for diarrhea or from patients first seeking medical
care from gastroenterologist. In order to minimize nosocomial infections
arising from unsuspected patients with diarrhea and gain comprehensive
understanding of transmission routes for this viral pathogen, they
compared the transcriptional levels of ACE2 of various human tissues
from NCBI public database as well as in small intestine tissue from
CD57BL/6 mice using single cell sequencing. They show that ACE2
expression is not only increased in the human small intestine, but
demonstrate a particular increase in mice enterocytes positioned on the
surface of the intestinal lining exposed to viral pathogens. Given that
ACE2 is the viral receptor for SARS-CoV-2 and also reported to regulate
diarrhea, their data suggests the small intestine as a potential
transmission route and diarrhea as a potentially underestimated symptom
in COVID19 patients that must be carefully monitored. Interestingly,
however, they show that ACE2 expression level is not elevated in human
lung tissue.
13.3.3 Limitations
Although this study demonstrates a
statistical difference in the incidence of diarrhea across three
separate COVID19 patient cohorts, their conclusions are limited by a
small sample size. Specifically, the p-value computed by Fisher’s exact
test is based on a single patient cohort of only six cases of which 33%
are reported to have diarrhea, while the remaining two larger cohorts
with 41 and 99 cases report 3% and 2% diarrhea incidence, respectively.
Despite showing significance, they would need to acquire larger sample
sizes and cohorts to minimize random variability and draw meaningful
conclusions. Furthermore, they do not address why ACE2 expression level
is not elevated in human lung tissue despite it being a major
established route of transmission for SARS-CoV-2. It could be helpful to
validate this result by looking at ACE2 expression in mouse lung tissue.
Finally, although this study is descriptive and shows elevated ACE2
expression in small intestinal epithelial cells, it does not establish a
mechanistic link to SARS-CoV-2 infection of the host. Overall, their
claim that infected patients exhibiting diarrhea pose an increased risk
to hospital staff needs to be further substantiated.
13.3.4 Significance
This study provides a possible transmission route and a
potentially underappreciated clinical symptom for SARS-CoV-2 for better
clinical management and control of COVID19.
13.3.5 Credit
Summary generated as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.4 Specific ACE2 Expression in Cholangiocytes May Cause Liver Damage After 2019-nCoV Infection
Using both publicly available scRNA-seq dataset of liver
samples from colorectal patients and scRNA-sequencing of four liver
samples from healthy volunteers, the authors show that ACE2 is
significantly enriched in the majority of cholangiocytes (59.7 %) but
not in hepatocytes (2.6%).
13.4.3 Main Findings
Using bioinformatics approaches of
RNASeq analysis, this study reveals that ACE2 dominates in
cholangiocytes and is present at very low levels in hepatocytes.
13.4.4 Limitations
The
study does not provide mechanistic insights into how SARS-CoV-2 can
infect and replicate in cholangiocytes and the types of intrinsic
anti-viral responses induced by cholangiocytes when infected. In
addition, because the study relies on the assumption that SARS-CoV-2
infects cells only through ACE2, it cannot discount the possibility that
the virus can infect hepatocytes through mechanisms other than
ACE2-mediated entry. Furthermore, because the scRNA-seq analysis were
performed on healthy liver samples, one cannot draw any definitive
conclusions about gene expression states (including ACE2 expression in
liver cell types) in system-wide inflammatory contexts.
13.4.5 Significance
This article with other studies on liver
damage in COVID patients suggests that liver damage observed in COVID
patients is more due to inflammatory cytokines than direct infection of
the liver. Even if cholangiocytes are infectable by SARS-CoV-2 (which
was demonstrated by human liver ductal organoid study
([1252]),
published clinical data show no significant increase in bile duct injury
related indexes (i.e. alkaline phosphatase, gamma-glutamyl
transpeptidase and total bilirubin). In sum, it underscores the
importance of future studies characterizing cellular responses of
extra-pulmonary organs in the context of COVID or at least in viral lung
infections..
13.4.6 Credit
Summary generated by Chang Moon as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.5 ACE2 expression by colonic epithelial cells is associated with viral infection, immunity and energy metabolism
Colonic enterocytes primarily express ACE2. Cellular pathways associated with ACE2 expression include innate immune signaling, HLA up regulation, energy metabolism and apoptotic signaling.
13.5.3 Limitations
This is a study of colonic biopsies taken from 17 children with and without IBD and analyzed using scRNAseq to look at ACE2 expression and identify gene families correlated with ACE2 expression. The authors find ACE2 expression to be primarily in colonocytes. It is not clear why both healthy and IBD patients were combined for the analysis. Biopsies were all of children so extrapolation to adults is limited. The majority of genes found to be negatively correlated with ACE2 expression include immunoglobulin genes (IGs). IG expression will almost certainly be low in colonocytes irrespective of ACE2 expression.
13.5.4 Significance
This study performs a retrospective analysis of ACE2 expression using an RNAseq dataset from intestinal biopsies of children with and without IBD. The implications for the CoV-19 epidemic are modest, but do provide support that ACE2 expression is specific to colonocytes in the intestines. The ontological pathway analysis provides some limited insights into gene expression associated with ACE2.
13.5.5 Credit
Summary generated as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.6 The Pathogenicity of 2019 Novel Coronavirus in hACE2 Transgenic Mice
Using a transgenic human Angiotensin-converting enzyme 2
(hACE2) mouse that has previously been shown susceptible to infection by
SARS-CoV, Bao et al. create a model of pandemic 2019-nCoV strain
coronavirus. The model includes interstitial hyperplasia in lung tissue,
moderate inflammation in bronchioles and blood vessels, and histology
consistent with viral pneumonia at 3 days post infection. Wildtype did
not experience these symptoms. In addition, viral antigen and hACE2
receptor were found to co-localize the lung by immunofluorescence 3-10
days post infection only in the hACE2 infected mice.
13.6.3 Limitations
The characterization of the infection remains incomplete,
as well as lacking characterization of the immune response other than
the presence of a single antiviral antibody. Though they claim to
fulfill Koch’s postulates, they only isolate the virus and re-infect
Vero cells, rather than naive mice.
13.6.4 Significance
This paper establishes a murine model for 2019-nCoV
infection with symptoms consistent with viral pneumonia. Though not
fully characterized, this model allows in vivo analysis of viral entry
and pathology that is important for the development of vaccines and
antiviral therapeutics.
13.6.5 Credit
Review by Dan Fu Ruan, Evan Cody and Venu Pothula as part of a project by students, postdocs and faculty
at the Immunology Institute of the Icahn School of Medicine, Mount
Sinai.
13.7 Caution on Kidney Dysfunctions of 2019-nCoV Patients
Retrospective study of 59 patients assayed key function indicators
of the kidney–including urine protein, blood urea nitrogen (BUN),
plasma creatinine (Cre), and renal CT scan data.
Found that 34% of patients developed massive albuminuria on the
first day of admission, and 63% developed proteinuria during their
stay in hospital; and 19% of patients had high plasma creatinine,
especially the terminal cases.
CT analyses of 27 patients showed all patients to have abnormal
kidney damage; indicate that inflammation and edema of the renal
parenchyma very common.
13.7.3 Limitations
No analysis of immunity-dependent damage and cytokines in
blood/plasma/urine. Will be worth correlating disease progression
with cytokine production, immune activity and kidney function.
Extrapolating to earlier SARS-CoV studies provides the only
rationale for viral-damage in kidney and resultant pathologic immune
response (understandable for this clinical study).
13.7.4 Significance
Multiple lines of evidence along this study’s finding point to the
idea that renal impairment/injury is a key risk factor in 2019-nCoV
patients similar to what has been reported for SARS-CoV[1256]; this
may be one of the major causes of virally-induced damage and
contribute to multiorgan failure.
ACE2 expression in kidney proximal tubule epithelia and bladder
epithelia [1257] support
these clinical findings.
Study argues for closely monitoring kidney function, and applying
potential interventions including continuous renal replacement
therapies (CRRT) for protecting kidney functions as early as
possible, particularly for those with rising plasma creatinine.
13.7.5 Credit
Review by Samarth Hegde as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.8 Profiling the immune vulnerability landscape of the 2019 Novel Coronavirus
This study harnesses bioinformatic profiling to predict the potential of
COV2 viral proteins to be presented on MHC I and II and to form linear
B-cell epitopes. These estimates suggest a T-cell antigenic profile
distinct from SARS-CoV or MERS-CoV, identify focused regions of the
virus with a high density of predicted epitopes, and provide preliminary
evidence for adaptive immune pressure in the genetic evolution of the
virus.
13.8.3 Limitations
While the study performs a comprehensive analysis of potential epitopes
within the virus genome, the analysis relies solely on bioinformatic
prediction to examine MHC binding affinity and B-cell epitope potential
and does not capture the immunogenicity or recognition of
these epitopes. Future experimental validation in data from patients
infected with SARS-CoV-2 will be important to validate and refine these
findings. Thus some of the potential conclusions stated, including viral
evolution toward lower immunogenicity or a dominant role for CD4+
T-cells rather than CD8+ T-cells in viral clearance, require further
valiadtion.
13.8.4 Significance
These findings may help direct peptide vaccine design toward relevant
epitopes and provide intriguing evidence of viral evolution in response
to immune pressure.
13.8.5 Credit
Summary generated as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.9 Single-cell Analysis of ACE2 Expression in Human Kidneys and Bladders Reveals a Potential Route of 2019-nCoV Infection
To investigate the possible cause of kidney damage in 2019-nCoV
patients, authors used published kidney and bladder cell atlas data
(GSE131685, GSE108097; 3 healthy donors each) as well as an
unpublished kidney single-cell RNA-Seq data (in-house from 2
transplant donors) to evaluate ACE2 gene expressions in all cell
types of healthy kidneys and bladders.
They find enriched expression of ACE2 transcript in all subtypes of
proximal tubule cells of kidney, with 5%-15% of both straight and
convoluted proximal tubule cells expressing ACE2.
They also find detectable levels of ACE2 in bladder epithelial
cells, noting expression from around 1.5% of cells in the outer
layer umbrella cells of the bladder epithelium and decreasing in the
basal cells.
Importantly endothelial or immune cells in kidney/bladder do not
express ACE2.
13.9.3 Limitations
This study primarily characterizes ACE2 expression (amongst other
genes) from a small healthy-donor dataset, and will benefit from
supporting data in (expired) patient samples to show functional
viral damage. ACE2 transcript does not necessarily translate to
viral permissiveness in kidney/bladder epithelia or cytokine
release.
This study focuses on only healthy tissue; it will be useful to
analyze kidney/bladder epithelial ACE2 expression under inflammatory
conditions or in patients with underlying kidney conditions.
Given what is known about protease TMPRSS2 expression during
SARS-CoV-2 infection, ACE2+TMPRSS2+ double-positive cell
identification would be useful in these datasets.
13.9.4 Significance
ACE2 protein is spatially restricted to brush border of proximal
tubules and in bladder umbrella cells [96], such cells in direct
contact with viral particles are likely to be highly sensitive to
viral-induced damage.
SARS-CoV and MERS-CoV have been shown to be detected in urine of
patients and associate with higher mortality [1256,1259], thus worth
understanding kidney damage and resultant immune response in
SARS-CoV-2 as well.
This study argues for a potential mode of viral infectivity and
resultant inflammatory responses in these tissue in addition to
reported infectivity in the lung and digestive system, which is
supported by clinical data showing acute and early kidney
complications in 2019-nCoV patients [1255].
Clinically, thus very important to track urinary CoVID-19 shedding
as well as study acute kidney injury-related co-morbidities.
13.9.5 Credit
Review by Samarth Hegde as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.10 Neutrophil-to-Lymphocyte Ratio Predicts Severe Illness Patients with 2019 Novel Coronavirus in the Early Stage
This study aimed to find prognostic biomarkers of COVID-19
pneumonia severity. Sixty-one (61) patients with COVID-19 treated in
January at a hospital in Beijing, China were included. On average,
patients were seen within 5 days from illness onset. Samples were
collected on admission; and then patients were monitored for the
development of severe illness with a median follow-up of 10 days].
Patients were grouped as “mild” (N=44) or “moderate/severe” (N=17)
according to symptoms on admission and compared for different
clinical/laboratory features. “Moderate/severe” patients were
significantly older (median of 56 years old, compared to 41 years old).
Whereas comorbidies rates were largely similar between the groups,
except for hypertension, which was more frequent in the severe group (p=
0.056). ‘Severe’ patients had higher counts of neutrophils, and serum
glucose levels; but lower lymphocyte counts, sodium and serum chlorine
levels. The ratio of neutrophils to lymphocytes (NLR) was also higher
for the ‘severe’ group. ‘Severe’ patients had a higher rate of bacterial
infections (and antibiotic treatment) and received more intensive
respiratory support and treatment.
26 clinical/laboratory variables were used to select NLR and age as the
best predictors of the severe disease. Predictive cutoffs for a severe
illness as NLR ≥ 3.13 or age ≥ 50 years.
13.10.3 Limitations
Identification of early biomarkers is important for
making clinical decisions, but large sample size and validation cohorts
are necessary to confirm findings. It is worth noting that patients
classified as “mild” showed pneumonia by imaging and fever, and in
accordance with current classifications this would be consistent with
“moderate” cases. Hence it would be more appropriate to refer to the
groups as “moderate” vs “severe/critical”. Furthermore, there are
several limitations that could impact the interpretation of the results:
e.g. classification of patients was based on symptoms presented on
admission and not based on disease progression, small sample size,
especially the number of ‘severe’ cases (with no deaths among these
patients). Given the small sample size, the proposed NLR and age cut
offs might not hold for a slightly different set of patients. For
example, in a study of >400 patients, ‘non-severe’ and ‘severe’ NLR
were 3.2 and 5.5, respectively [1261].
13.10.4 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.11 Characteristics of lymphocyte subsets and cytokines in peripheral blood of 123 hospitalized patients with 2019 novel coronavirus pneumonia (NCP)
The authors analyzed lymphocyte subsets and cytokines of 102 patients
with mild disease and 21 with severe disease. CD8+T cells and CD4+T
cells were significantly reduced in both cohort. particularly in severe
patients. The cytokines IL6 and IL10 were significantly elevated in
severe patients as compared to mild. No significant differences were
observed in frequency of B cells and NK cells.
The authors argue that the measurement of T cell frequencies and
cytokine levels of IL6 and IL10 can be used to predict progression of
disease from Mild to severe Cov-2 infection.
13.11.3 Limitations
The study demonstrates in a limited cohort
similar associations to several other reported studies. The authors
didn’t compare the changes in lymphocyte and cytokine with healthy
individual (Covid-19 Negative) rather used an internal standard value.
The recently preprint in LANCET shows The degree of lymphopenia and a
pro-inflammatory cytokine storm is higher in severe COVID-19 patients
than in mild cases, and is associated with the disease severity [1263].
13.11.4 Significance
This translational data identifies key cytokines and lymphopenia
associated with disease severity although mechanism and key cellular
players are still unknown. Higher level IL-6 production in severe
patient suggests potential role of Tocilizumab (anti-IL6R) biologic
although clinical trial will be necessary.
13.11.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.12 Epidemiological and Clinical Characteristics of 17 Hospitalized Patients with 2019 Novel Coronavirus Infections Outside Wuhan, China
These authors looked at 17 hospitalized patients with COVID-19
confirmed by RT-PCR in Dazhou, Sichuan. Patients were admitted between
January 22 and February 10 and the final data were collected on February
11. Of the 17 patients, 12 remained hospitalized while 5 were discharged
after meeting national standards. The authors observed no differences
based on the sex of the patients but found that the discharged patients
were younger in age (p = 0.026) and had higher lymphocyte counts (p =
0.005) and monocyte counts (p = 0.019) upon admission.
13.12.3 Limitations
This study is limited in the sample size of the study and the last
data collection point was only one day after some of the patients were
admitted.
13.12.4 Significance
These findings have been somewhat supported by subsequent studies
that show that older age and an immunocompromised state are more likely
to result in a more severe clinical course with COVID-19. However, other
studies have been published that report on larger numbers of cases.
13.12.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.13 ACE2 Expression in Kidney and Testis May Cause Kidney and Testis Damage After 2019-nCoV Infection
Study used online datasets (scRNAseq GSE131685, scRNAseq GSE107585,
Human Protein Atlas, GTEx portal, CCLE) to analyze ACE2 expression
in different human organs.
Study re-analyzed three clinical datasets (n=6, n=99, and n=41) to
show 3~10% of 2019-nCoV patients present with abnormal renal
function.
results indicate ACE2 highly expressed in renal tubular cells,
Leydig cells and seminiferous ductal cells of testis.
13.13.3 Limitations
Very preliminary transcript/protein dataset analysis in healthy
cohorts; does not necessarily translate to actual viral tropism and
permissiveness.
Clinically, would be important to determine with larger longitudinal
dataset if SARS-CoV-2 infection changes sperm quality or testicular
inflammation.
Similarly, would be important to determine if simultaneous HBV or
syphilis infection and orchitis impacts SARS-CoV-2 severity.
Examination and follow-up of renal function and viral orchitis/sperm
quality of CoVID-19 patients not done in this preliminary study.
13.13.4 Significance
Kidney ACE2 result supports other concurrent sequencing studies
[1257] and clinical reports
of abnormal renal function or even kidney damage in patients
infected with 2019-nCoV
[1255].
High ACE2 expression in testis suggests potential tropism of the
virus to testicular tissues and indicates potential risks for male
fertility. Viral orchitis reported for SARS-CoV previously [1],
but no clear evidence so far of infertility in SARS, MERS or
CoVID-19 patients.
13.13.5 Credit
Review by Samarth Hegde as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.14 Aberrant pathogenic GM-CSF+ T cells and inflammatory CD14+CD16+ monocytes in severe pulmonary syndrome patients of a new coronavirus
The authors of this study sought to characterize the immune
mechanism causing severe pulmonary disease and mortality in 2019-nCoV
(COVID-19) patients. Peripheral blood was collected from hospitalized
ICU (n=12) and non-ICU (n=21) patients with confirmed 2019-nCoV and from
healthy controls (n=10) in The First Affiliated Hospital of University
of Science and Technology China (Hefei, Anhui). Immune analysis was
conducted by flow cytometry. 2019-nCoV patients had decreased
lymphocyte, monocyte, and CD4 T cell counts compared to healthy
controls. ICU patients had fewer lymphocytes than non-ICU patients. CD4
T cells of 2019-nCoV patients expressed higher levels of activation
markers (OX40, CD69, CD38, CD44) and exhaustion markers (PD-1 and Tim3)
than those of healthy controls. CD4 cells of ICU patients expressed
significantly higher levels of OX40, PD-1, and Tim3 than those of
non-ICU patients. 2019-nCoV patients had higher percentages of CD4 T
cells co-expressing GM-CSF and IL-6 compared to healthy controls, while
ICU patients had a markedly higher percentage of GM-CSF+ IFN-γ+ CD4 T
cells than non-ICU patients. The CD4 T cells of nCoV patients and
healthy controls showed no differences in TNF-α secretion.
The CD8 T cells of 2019-nCoV patients also showed higher expression of
activation markers CD69, CD38, and CD44, as well as exhaustion markers
PD-1 and Tim3, compared to healthy controls. CD8 T cells of ICU patients
expressed higher levels of GM-CSF than those of non-ICU patients and
healthy controls. No IL-6 or TNF-α was found in the CD8 T cells of any
group. There were no differences in numbers of NK cells or B cells in
2019-nCoV patients and healthy controls, nor was there any GM-CSF or
IL-6 secretion from these cells in either group.
Percentages of CD14+ CD16+ GM-CSF+ and CD14+ CD16+ IL-6+ inflammatory
monocytes were significantly increased in nCoV patients compared to
healthy controls; in particular, patients in the ICU had greater
percentages of CD14+ CD16+ IL-6+ monocytes than non-ICU patients. The
authors suggest that in 2019-nCoV patients, pathogenic Th1 cells produce
GM-CSF, recruiting CD14+ CD16+ inflammatory monocytes that secrete high
levels of IL-6. These may enter pulmonary circulation and damage lung
tissue while initiating the cytokine storm that causes mortality in
severe cases. This is consistent with the cytokine storm seen in similar
coronaviruses, as IL-6, IFN-γ, and GM-CSF are key inflammatory mediators
seen in patients with SARS-CoV-1 and MERS-CoV.
13.14.3 Limitations
Though the results of this study open
questions for further investigation, this is an early study on a small
cohort of patients, and as such there are a number of limitations. The
study included only 12 ICU patients and 21 non-ICU patients, and ideally
would be repeated with a much larger patient cohort. Though the authors
make claims about differences in lymphocyte and monocyte counts between
patients and healthy controls, they did not report baseline laboratory
findings for the control group. Additionally, severity of disease was
classified based on whether or not patients were in the ICU. It would be
interesting to contextualize the authors’ immunological findings with
more specific metrics of disease severity or time course. Noting
mortality, time from disease onset, pre-existing conditions, or severity
of lung pathology in post-mortem tissue samples would paint a fuller
picture of how to assess risk level and the relationship between
severity of disease and immunopathology. Another limitation is the
selection of cytokines and immune markers for analysis, as the selection
criteria were based on the cell subsets and cytokine storm typically
seen in SARS-CoV-1 and MERS-CoV patients. Unbiased cytokine screens and
immune profiling may reveal novel therapeutic targets that were not
included in this study.
13.14.4 Significance
This study identifies potential therapeutic targets
that could prevent acute respiratory disease syndrome (ARDS) and
mortality in patients most severely affected by COVID-19. The authors
propose testing monoclonal antibodies against IL6-R or GM-CSF to block
recruitment of inflammatory monocytes and the subsequent cytokine storm
in these patients.
13.14.5 Credit
Review by Gabrielle Lubitz as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.15 Clinical Characteristics of 2019 Novel Infected Coronavirus Pneumonia:A Systemic Review and Meta-analysis
The authors performed a meta analysis of literature on clinical,
laboratory and radiologic characteristics of patients presenting with
pneumonia related to SARSCoV2 infection, published up to Feb 6 2020.
They found that symptoms that were mostly consistent among studies were
sore throat, headache, diarrhea and rhinorrhea. Fever, cough, malaise
and muscle pain were highly variable across studies. Leukopenia (mostly
lymphocytopenia) and increased white blood cells were highly variable
across studies. They identified three most common patterns seen on CT
scan, but there was high variability across studies. Consistently across
the studies examined, the authors found that about 75% of patients need
supplemental oxygen therapy, about 23% mechanical ventilation and about
5% extracorporeal membrane oxygenation (ECMO). The authors calculated a
staggering pooled mortality incidence of 78% for these patients.
13.15.3 Limitations
The authors mention that the total number of studies included in this
meta analysis is nine, however they also mentioned that only three
studies reported individual patient data. It is overall unclear how many
patients in total were included in their analysis. This is mostly
relevant as they reported an incredibly high mortality (78%) and mention
an absolute number of deaths of 26 cases overall. It is not clear from
their report how the mortality rate was calculated.
The data is based on reports from China and mostly from the Wuhan area,
which somewhat limits the overall generalizability and applicability of
these results.
13.15.4 Significance
This meta analysis offers some important data for clinicians to refer to
when dealing with patients with COVID-19 and specifically with
pneumonia. It is very helpful to set expectations about the course of
the disease.
13.15.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.16 Longitudinal characteristics of lymphocyte responses and cytokine profiles in the peripheral blood of SARS-CoV-2 infected patients
Liu et al. enrolled a cohort of 40 patients from Wuhan
including 27 mild cases and 13 severe cases of COVID-19. They performed
a 16-day kinetic analysis of peripheral blood from time of disease
onset. Patients in the severe group were older (medium age of 59.7,
compared to 48.7 in mild group) and more likely to have hypertension as
a co-morbidity. Lymphopenia was observed in 44.4% of the mild patients
and 84.6% of the severe patients. Lymphopenia was due to low T cell
count, specially CD8 T cells. Severe patients showed higher neutrophil
counts and an increase of cytokines in the serum (IL2, IL6, IL10 and
IFNγ). The authors measured several other clinical laboratory parameters
were also higher in severe cases compared to mild, but concluded that
neutrophil to CD8 T cell ratio (N8R) as the best prognostic factor to
identify the severe cases compared to other receiver operating
characteristic (ROC).
13.16.3 Limitations
This was a small cohort (N=40), and two of the patients
initially included in the severe group (N=13) passed away and were
excluded from the analysis due to lack of longitudinal data. However, it
would be most important to be able to identify patients with severe
disease with higher odds of dying. It seems that the different time
points analyzed relate to hospital admission, which the authors describe
as disease onset. The time between first symptoms and first data points
is not described. It would have been important to analyze how the
different measured parameters change according to health condition, and
not just time (but that would require a larger cohort). The predictive
value of N8R compared to the more commonly used NLR needs to be assessed
in other independent and larger cohorts. Lastly, it is important to note
that pneumonia was detected in patients included in the “mild” group,
but according to the Chinese Clinical Guidance for COVID-19 Pneumonia
Diagnosis and Treatment (7th edition) this group should be considered
“moderate”.
13.16.4 Significance
Lymphopenia and cytokine storm have been described to be
detrimental in many other infections including SARS-CoV1 and MERS-CoV.
However, it was necessary to confirm that this dramatic immune response
was also observed in the SARS-CoV2 infected patients. These results and
further validation of the N8R ratio as a predictor of disease severity
will contribute for the management of COVID19 patients and potential
development of therapies.
13.16.5 Credit
Review by Pauline Hamon as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.17 Clinical and immunologic features in severe and moderate forms of Coronavirus Disease 2019
This study retrospectively evaluated clinical, laboratory,
hematological, biochemical and immunologic data from 21 subjects
admitted to the hospital in Wuhan, China (late December/January) with
confirmed SARS-CoV-2 infection. The aim of the study was to compare
‘severe’ (n=11, ~64 years old) and ‘moderate’ (n=10, ~51 years old)
COVID-19 cases. Disease severity was defined by patients’ blood oxygen
level and respiratory output. They were classified as ‘severe’ if SpO2
93% or respiratory rates 30 per min.
In terms of the clinical laboratory measures, ‘severe’ patients had
higher CRP and ferritin, alanine and aspartate aminotransferases, and
lactate dehydrogenase but lower albumin concentrations.
The authors then compared plasma cytokine levels (ELISA) and immune cell
populations (PBMCs, Flow Cytometry). ‘Severe’ cases had higher levels of
IL-2R, IL-10, TNFa, and IL-6 (marginally significant). For the immune
cell counts, ‘severe’ group had higher neutrophils, HLA-DR+ CD8 T cells
and total B cells; and lower total lymphocytes, CD4 and CD8 T cells
(except for HLA-DR+), CD45RA Tregs, and IFNy-expressing CD4 T cells. No
significant differences were observed for IL-8, counts of NK cells,
CD45+RO Tregs, IFNy-expressing CD8 T and NK cells.
13.17.3 Limitations
Several potential limitations should be noted: 1) Blood samples were
collected 2 days post hospital admission and no data on viral loads were
available; 2) Most patients were administered medications (e.g.
corticosteroids), which could have affected lymphocyte counts.
Medications are briefly mentioned in the text of the manuscript; authors
should include medications as part of Table 1. 3) ‘Severe’ cases were
significantly older and 4/11 ‘severe’ patients died within 20 days.
Authors should consider a sensitivity analysis of biomarkers with the
adjustment for patients’ age.
13.17.4 Significance
Although the sample size was small, this paper presented a broad range
of clinical, biochemical, and immunologic data on patients with
COVID-19. One of the main findings is that SARS-CoV-2 may affect T
lymphocytes, primarily CD4+ T cells, resulting in decreased IFNy
production. Potentially, diminished T lymphocytes and elevated cytokines
can serve as biomarkers of severity of COVID-19.
13.17.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.18 SARS-CoV-2 and SARS-CoV Spike-RBD Structure and Receptor Binding Comparison and Potential Implications on Neutralizing Antibody and Vaccine Development
This study compared the structure of SARS-CoV and SARS-CoV-2 Spike (S)
protein receptor binding domain (RBD) and interactions with ACE2 using
computational modeling, and interrogated cross-reactivity and
cross-neutralization of SARS-CoV-2 by antibodies against SARS-CoV. While
SARS-CoV and SARS-CoV-2 have over 70 % sequence homology and share the
same human receptor ACE2, the receptor binding motif (RBM) is only 50%
homologous.
Computational prediction of the SARS-CoV-2 and ACE2 interactions based
on the previous crystal structure data of SARS-CoV, and measurement of
binding affinities against human ACE2 using recombinant SARS-CoV and
SARS-CoV-2 S1 peptides, demonstrated similar binding of the two S1
peptides to ACE2, explaining the similar transmissibility of SARS-CoV
and SARS-CoV-2 and consistent with previous data (Wall et al Cell
2020).
The neutralization activity of SARS-CoV-specific rabbit polyclonal
antibodies were about two-order of magnitude less efficient to
neutralize SARS-CoV-2 than SARS-CoV, and four potently neutralizing
monoclonal antibodies against SARS-CoV had poor binding and neutralizing
activity against SARS-CoV-2. In contrast, 3 poor SARS-CoV-binding
monoclonal antibodies show some efficiency to bind and neutralize
SARS-CoV-2. The results suggest that that antibodies to more conserved
regions outside the RBM motif might possess better cross-protective
neutralizing activities between two strains.
13.18.3 Limitations
It would have been helpful to show the epitopes recognized by the
monoclonal antibodies tested on both SARS-CoV, SARS-CoV-2 to be able to
make predictions for induction of broadly neutralizing antibodies. The
data on monoclonal antibody competition with ACE2 for binding to
SARS-CoV RBD should have also included binding on SARS-CoV2, especially
for the three monoclonal antibodies that showed neutralization activity
for SARS-CoV2. Because of the less homology in RBM sequences between
viruses, it still may be possible that these antibodies would recognize
the ACE2 RBD in SARS-CoV-2.
13.18.4 Significance
It is noteworthy that immunization to mice and rabbit with SARS-CoV S1
or RBD protein could induce monoclonal antibodies to cross-bind and
cross-neutralize SARS-CoV-2 even if they are not ACE2-blocking. If these
types of antibodies could be found in human survivors or in the
asymptomatic populations as well, it might suggest that exposure to
previous Coronavirus strains could have induced cross-neutralizing
antibodies and resulted in the protection from severe symptoms in some
cases of SARS-CoV2.
13.18.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.19 Protection of Rhesus Macaque from SARS-Coronavirus challenge by recombinant adenovirus vaccine
Rhesus macaques were immunized intramuscularly twice (week 0 and
week 4) with SV8000 carrying the information to express a S1-orf8
fusion protein and the N protein from the BJ01 strain of SARS-CoV-1.
By week 8, immunized animals had signs of immunological protection
(IgG and neutralization titers) against SARS-CoV-1 and were
protected against challenge with the PUMC-1 strain, with fewer
detectable symptoms of respiratory distress, lower viral load,
shorter periods of viral persistence, and less pathology in the
lungs compared to non-immunized animals.
13.19.3 Limitations
The authors should write clearer descriptions of the methods used in
this article. They do not describe how the IgG titers or
neutralization titers were determined. There are some issues with
the presentation of data, for example, in Figure 1a, y-axis should
not be Vmax; forming cells and 1d would benefit from showing error
bars. Furthermore, although I inferred that the animals were
challenged at week 8, the authors did not explicitly detail when the
animals were challenged. The authors should explain the design of
their vaccine, including the choice of antigens and vector. The
authors also do not include a description of the ethical use of
animals in their study.
13.19.4 Significance
The authors describe a vaccine for SARS-CoV-1 with no discussion of
possible implications for the current SARS-CoV-2 pandemic. Could a
similar vaccine be designed to protect against SARS-CoV-2 and would
the concerns regarding emerging viral mutations that the authors
describe as a limitation for SARS-CoV-1 also be true in the context
of SARS-CoV-2?
13.19.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.20 Reduction and Functional Exhaustion of T cells in Patients with Coronavirus Disease 2019 (COVID-19)
Based on a retrospective study of 522 COVID patients and 40
healthy controls from two hospitals in Wuhan, China, authors show both
age-dependent and clinical severity-dependent decrease in T cell numbers
with elderly patients and patients who are in ICU-care showing the most
dramatic decrease in T cell counts. Cytokine profiling of COVID patients
reveal that TNF-α, IL-6 and IL-10 are increased in infected patients
with patients in the ICU showing the highest levels. Interestingly,
these three cytokine levels were inversely correlated with T cell counts
and such inverse relationship was preserved throughout the disease
progression. Surface staining of exhaustion markers (PD-1 and Tim-3) and
flow cytometry of stained peripheral blood of 14 patients and 3 healthy
volunteers demonstrate that T cells of COVID patients have increased
expression of PD-1 with patients in ICU having the highest number of
CD8+PD-1+ cells than their counterparts in non-ICU groups.
13.20.3 Limitations
Compared to the number of patients, number of control (n=
40) is small and is not controlled for age. Additional data linking
inflammatory cytokines and the quality of the adaptive response
including humoral and antigen specific T cell response is much needed. T
cell exhaustion study relies on marker-dependent labeling of T cell
functionality of a very limited sample size (n=17)—a
functional/mechanistic study of these T cells from PBMCs would have
bolstered their claims.
13.20.4 Significance
Limited but contains interesting
implications. It is already known in literature that in the context of
acute respiratory viral infections CD8 T cells exhibit exhaustion-like
phenotypes which further underscores the importance of mechanistic
studies that can elucidate how COVID infection leads to lymphopenia and
T cell exhaustion-like phenotype.
However, as authors have noted, the data does point to an interesting
question: How these inflammatory cytokines (TNF-α, IL-6 and IL-10)
correlate with or affect effective viral immunity and what types of
cells produce these cytokines? Answering that question will help us
refine our targets for immune-modulatory therapies especially in
patients suffering from cytokine storms.
13.20.5 Credit
This review by Chang Moon was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.21 Clinical Characteristics of 25 death cases infected with COVID-19 pneumonia: a retrospective review of medical records in a single medical center, Wuhan, China
Most common chronic conditions among 25 patients that died from
COVID-19 related respiratory failure were hypertension (64%) and
diabetes (40%). Disease progression was marked by progressive organ
failure, starting first with lung dysfunction, then heart (e.g.
increased cTnI and pro-BNP), followed by kidney (e.g. increased BUN,
Cr), and liver (e.g. ALT, AST). 72% of patients had neutrophilia and 88%
also had lymphopenia. General markers of inflammation were also
increased (e.g. PCT, D-Dimer, CRP, LDH, and SAA).
13.21.3 Limitations
The limitations of this study include small sample size and
lack of measurements for some tests for several patients. This study
would also have been stronger with comparison of the same measurements
to patients suffering from less severe disease to further validate and
correlate proposed biomarkers with disease severity.
13.21.4 Significance
This study identifies chronic conditions (i.e. hypertension
and diabetes) that strongly correlates with disease severity. In
addition to general markers of inflammation, the authors also identify
concomitant neutrophilia and lymphopenia among their cohort of patients.
This is a potentially interesting immunological finding because we would
typically expect increased lymphocytes during a viral infection.
Neutrophilia may also be contributing to cytokine storm. In addition,
PCT was elevated in 90.5% of patients, suggesting a role for sepsis or
secondary bacterial infection in COVID-19 related respiratory failure.
13.21.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.22 SARS-CoV-2 infection does not significantly cause acute renal injury: an analysis of 116 hospitalized patients with COVID-19 in a single hospital, Wuhan, China
Clinical data from 116 hospitalized CoVID-19 patients analyzed over
4 weeks for correlation with renal injury. Comorbidities included
chronic renal failure (CRF) in 5 patients (4.3%).
10.8% of patients with no prior kidney disease showed elevations in
blood urea or creatinine, and 7.2% of patients with no prior kidney
disease showed albuminuria.
Patients with pre-existing CRF underwent continuous renal
replacement therapy (CRRT) alongside CoVID-19 treatment. Renal
functions remained stable in these patients.
All 5 patients with CRF survived CoVID-19 therapy without
progression to ARDS or worsening of CRF.
13.22.3 Limitations
Renal injury biomarkers in patients with incipient kidney
abnormalities not tabulated separately, making overall data hard to
interpret. It will be critical to separately examine kidney function
(BUN, urine creatinine and eGFR) in patients that developed any
kidney abnormalities (7.2~10.8% of cohort).
No information on type of CoVID-19 therapy used across cohort; will
be useful to correlate how treatment modality influences kidney
function (and other parameters).
Invokes previous clinical-correlation studies that indicate low
instances of kidney damage[1274,1275], but those studies did not track
longitudinal urine samples for acute renal injury markers and viral
shedding.
CRRT in patients with CRF is standard therapy irrespective of
CoVID-19 status; it will be important to compare clinical parameters
of these patients (n=5) with virus-naïve CRF patients (none in this
study) to make any meaningful conclusions.
13.22.4 Significance
This study argues that renal impairment is uncommon in CoVID-19 and
not associated with high mortaility, in stark contrast with a
concurrent study [1255].
If supported by further studies, this argues kidney impairment is
secondary to cytokine storm/inflammation-induced organ failure, and
not due to direct viral replication.
Will be important to comprehensively characterize large-datasets of
CoVID-19 patients to conclude if kidney function actively disrupted
due to viral infection.
13.22.5 Credit
Review by Samarth Hegde as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.23 Potential T-cell and B-Cell Epitopes of 2019-nCoV
The authors use 2 neural network algorithms, NetMHCpan4 and
MARIA, to identify regions within the COVID-19 genome that are
presentable by HLA. They identify 405 viral epitopes that are
presentable on MHC-I and MHC-II and validate using known epitopes from
SARS-CoV. To determine whether immune surveillance drives viral
mutations to evade MHC presentation, the authors analyzed 68 viral
genomes from 4 continents. They identified 93 point mutations that
occurred preferentially in regions predicted to be presented by MHC-I
(p=0.02) suggesting viral evolution to evade CD8 T-cell mediated
killing. 2 nonsense mutations were also identified that resulted in loss
of presentation of an associated antigen (FGDSVEEVL) predicted to be
good antigen for presentation across multiple HLA alleles.
To identify potential sites of neutralizing antibody binding, the
authors used homology modeling to the SARS-CoV’s spike protein (S
protein) to determine the putative structure of the CoV2 spike protein.
They used Discotope2 to identify antibody binding sites on the protein
surface in both the down and up conformations of the S protein. The
authors validate this approach by first identifying antibody binding
site in SARS-CoV S protein. In both the down and up conformation of the
CoV2 S protein, the authors identified a potential antibody binding site
on the S protein receptor binding domain (RBD) of the ACE2 receptor
(residues 440-460, 494-506). While RBDs in both SARS-CoV and CoV2 spike
proteins may be important for antibody binding, the authors note that
SARS-CoV has larger attack surfaces than CoV2. These results were later
validated on published crystal structures of the CoV2 S protein RBD and
human ACE2. Furthermore, analysis of 68 viral genomes did not identify
any mutations in this potential antibody binding site in CoV2.
Finally, the authors compile a list of potential peptide vaccine
candidates across the viral genome that can be presented by multiple HLA
alleles. Several of the peptides showed homology to SARS-CoV T-cell and
B-cell epitopes.
13.23.3 Limitations
While the authors used computational methods of validation,
primarily through multiple comparisons to published SARS-CoV structures
and epitopes, future work should include experimental validation of
putative T-cell and B-cell epitopes.
13.23.4 Significance
The authors identified potential T-cell and B-cell epitopes
that may be good candidates for peptide based vaccines against CoV2.
They also made interesting observations in comparing SARS-CoV and CoV2
potential antibody binding sites, noting that SARS-CoV had larger attack
surfaces for potential neutralizing antibody binding. One of the
highlights of this paper was the authors’ mutation analysis of 68 viral
genomes from 4 continents. This analysis not only validated their
computational method for identifying T-cell epitopes, but showed that
immune surveillance likely drives viral mutation in MHC-I binding
peptides. The smaller attack surface may point to potential mechanisms
of immune evasion by CoV2. However, absence of mutations in the RBD of
CoV2 and the small number of mutations in peptides presentable to T
cells suggests that vaccines against multiple epitopes could still
elicit robust immunity against CoV2.
13.23.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.24 Structure, Function, and Antigenicity of the SARSCoV-2 Spike Glycoprotein
The authors highlight a human angiotensin-converting enzyme 2
(hACE2), as a potential receptor used by the current Severe Acute
respiratory syndrome coronavirus-2 (SARS-CoV-2) as a host factor that
allows the virus target human cells. This virus-host interaction
facilitates the infection of human cells with a high affinity comparable
with SARS-CoV. The authors propose this mechanism as a probable
explanation of the efficient transmission of SARS-CoV-2 between humans.
Besides, Walls and colleagues described SARS-CoV-2 S glycoprotein S by
Cryo-EM along with neutralizing polyclonal response against SAR-CoV-2 S
from mice immunized with SAR-CoV and blocking SAR-CoV-2 S-mediated entry
into VeroE6 infected cells.**
13.24.3 Limitations
The SARS-CoV-2 depends on the cell factors ACE2 and TMPRSS2, this
last, according to a recent manuscript by Markus Hoffman et al., Cell,
2020. The authors used green monkey (VeroE6) and hamster (BHK) cell
lines in the experiments to drive its conclusions to humans; however, it
is well known the caucasian colon adenocarcinoma human cell line
(CaCo-2), highly express the hACE2 receptor as the TMPRSS2 protease as
well. In humans, ACE2 protein is highly expressed in the
gastrointestinal tract, which again, makes the CaCo-2 cell line suitable
for the following SARS-CoV-2 studies.
13.24.4 Significance
The results propose a functional receptor used by SARS-CoV-2 to
infect humans worldwide and defining two distinct conformations of spike
(S) glycoprotein by cryogenic electron microscopy (Cryo-EM). This study
might help establish a precedent for initial drug design and treatment
of the current global human coronavirus epidemic.
13.24.5 Credit
Review by postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.25 Breadth of concomitant immune responses underpinning viral clearance and patient recovery in a non-severe case of COVID-19
The authors characterized the immune response in peripheral blood of a
47-year old COVID-19 patient.
SARS-CoV2 was detected in nasopharyngeal swab, sputum and faeces
samples, but not in urine, rectal swab, whole blood or throat swab. 7
days after symptom onset, the nasopharyngeal swab test turned negative,
at day 10 the radiography infiltrates were cleared and at day 13 the
patient became asymptomatic.
Immunofluorescence staining shows from day 7 the presence of
COVID-19-binding IgG and IgM antibodies in plasma, that increase
until day 20.
Flow cytometry on whole blood reveals a plasmablast peak at day 8, a
gradual increase in T follicular helper cells, stable HLA-DR+ NK
frequencies and decreased monocyte frequencies compared to healthy
counterparts. The expression of CD38 and HLA-DR peaked on T cells at D9
and was associated with higher production of cytotoxic mediators by
CD8+ T cells.
IL-6 and IL-8 were undetectable in plasma.
The authors further highlight the presence of the IFITM3
SNP-rs12252-C/C variant in this patient, which is associated with
higher susceptibility to influenza virus.
13.25.3 Limitations
These results need to be confirmed in additional patients.
COVID-19 patients have increased infiltration of macrophages in their
lungs [1279]. Monitoring monocyte proportions in blood earlier in the
disease might help to evaluate their eventual migration to the lungs.
The stable concentration of HLA-DR+ NK cells in blood from day 7 is
not sufficient to rule out NK cell activation upon SARS-CoV2 infection.
In response to influenza A virus, NK cells express higher levels of
activation markers CD69 and CD38, proliferate better and display higher
cytotoxicity [1280]. Assessing these parameters in COVID-19 patients is
required to better understand NK cell role in clearing this infection.
Neutralization potential of the COVID-19-binding IgG and IgM antibodies
should be assessed in future studies.
This patient was able to clear the virus, while presenting a SNP
associated with severe outcome following influenza infection. The
association between this SNP and outcome upon SARS-CoV2 infection should
be further investigated.
13.25.4 Significance
This study is among the first to describe the appearance of
COVID-19-binding IgG and IgM antibodies upon infection. The emergence of
new serological assays might contribute to monitor more precisely the
seroconversion kinetics of COVID-19 patients [392]. Further association
studies between IFITM3 SNP-rs12252-C/C variant and clinical data might
help to refine the COVID-19 outcome prediction tools.
13.25.5 Credit
Review by Bérengère Salomé as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.26 The landscape of lung bronchoalveolar immune cells in COVID-19 revealed by single-cell RNA sequencing
The authors performed single-cell RNA sequencing (scRNAseq) on
bronchoalveolar lavage fluid (BAL) from 6 COVID-19 patients (n=3 mild
cases, n=3 severe cases). Data was compared to previously generated
scRNAseq data from healthy donor lung tissue (n=8).
Clustering analysis of the 6 patients revealed distinct immune cell
organization between mild and severe disease. Specifically, they found
that transcriptional clusters annotated as tissue resident alveolar
macrophages were strongly reduced while monocytes-derived FCN1+SPP1+
inflammatory macrophages dominated the BAL of patients with severe
COVID19 diseases. They show that inflammatory macrophages upregulated
interferon-signaling genes, monocytes recruiting chemokines including
CCL2, CCL3, CCL4 as well as IL-6, TNF, IL-8 and profibrotic cytokine
TGF-β, while alveolar macrophages expressed lipid metabolism genes, such
as PPARG.
The lymphoid compartment was overall enriched in lungs from patients.
Clonally expanded CD8 T cells were enriched in mild cases suggesting
that CD8 T cells contribute to viral clearance as in Flu infection,
whereas proliferating T cells were enriched in severe cases.
SARS-CoV-2 viral transcripts were detected in severe patients, but
considered here as ambient contaminations.
13.26.3 Limitations
These results are based on samples from 6 patients and should therefore
be confirmed in the future in additional patients. Longitudinal
monitoring of BAL during disease progression or resolution would have
been most useful.
The mechanisms underlying the skewing of the macrophage compartment in
patients towards inflammatory macrophages should be investigated in
future studies.
Deeper characterization of the lymphoid subsets is required. The
composition of the “proliferating” cluster and how these cells differ
from conventional T cell clusters should be assessed. NK and CD8 T cell
transcriptomic profile, in particular the expression of cytotoxic
mediator and immune checkpoint transcripts, should be compared between
healthy and diseased lesions.
13.26.4 Significance
COVID-19 induces a robust inflammatory cytokine storm in patients that
contributes to severe lung tissue damage and ARDS [1281]. Accumulation of
monocyte-derived inflammatory macrophages at the expense of Alveolar
macrophages known to play an anti-inflammatory role following
respiratory viral infection, in part through the PPARγ pathway [1282,1283] are
likely contributing to lung tissue injuries. These data suggest that
reduction of monocyte accumulation in the lung tissues could help
modulate COVID-19-induced inflammation. Further analysis of lymphoid
subsets is required to understand the contribution of adaptive immunity
to disease outcome.
13.26.5 Credit
Review by Bérengère Salomé and Assaf Magen as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.27 Can routine laboratory tests discriminate 2019 novel coronavirus infected pneumonia from other community-acquired pneumonia?
In an attempt to use standard laboratory testing for the discrimination
between “Novel Coronavirus Infected Pneumonia” (NCIP) and a usual
community acquired pneumonia (CAP), the authors compared laboratory
testing results of 84 NCIP patients with those of a historical group of
316 CAP patients from 2018 naturally COVID-19 negative. The authors
describe significantly lower white blood- as well as red blood- and
platelet counts in NCIP patients. When analyzing differential blood
counts, lower absolute counts were measured in all subsets of NCIP
patients. With regard to clinical chemistry parameters, they found
increased AST and bilirubin in NCIP patients as compared to CAP
patients.
13.27.3 Limitations
The authors claim to describe a simple method to rapidly assess a
pre-test probability for NCIP. However, the study has substantial
weakpoints. The deviation in clinical laboratory values in NCIP patients
described here can usually be observed in severely ill patients. The
authors do not comment on how severely ill the patients tested here were
in comparison to the historical control. Thus, the conclusion that the
tests discriminate between CAP and NCIP lacks justification.
13.27.4 Significance
The article strives to compare initial laboratory testing results in
patients with COVID-19 pneumonia as compared to patients with a usual
community acquired pneumonia. The implications of this study for the
current clinical situation seem restricted due to a lack in clinical
information and the use of a control group that might not be
appropriate.
13.27.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.28 Correlation Analysis Between Disease Severity and Inflammation-related Parameters in Patients with COVID-19 Pneumonia
This study is a cross-sectional analysis of 100 patients with
COVID-19 pneumonia, divided into mild (n = 34), severe (n = 34), and
critical (n = 32) disease status based on clinical definitions.
The criteria used to define disease severity are as follows:
Severe – any of the following: respiratory distress or respiratory
rate ≥ 30 respirations/minute; oxygen saturation ≤ 93% at rest;
oxygen partial pressure (PaO2)/oxygen concentration (FiO2) in
arterial blood ≤ 300mmHg, progression of disease on imaging
to >50% lung involvement in the short term.
Critical – any of the following: respiratory failure that requires
mechanical ventilation; shock; other organ failure that requires
treatment in the ICU.
Patients with pneumonia who test positive for COVID-19 who do not
have the symptoms delineated above are considered mild.
Peripheral blood inflammatory markers were correlated to disease status.
Disease severity was significantly associated with levels of IL-2R,
IL-6, IL-8, IL-10, TNF-α, CRP, ferroprotein, and procalcitonin. Total
WBC count, lymphocyte count, neutrophil count, and eosinophil count were
also significantly correlated with disease status. Since this is a
retrospective, cross-sectional study of clinical laboratory values,
these data may be extrapolated for clinical decision making, but without
studies of underlying cellular causes of these changes this study does
not contribute to a deeper understanding of SARS-CoV-2 interactions with
the immune system.
It is also notable that the mean age of patients in the mild group was
significantly different from the mean ages of patients designated as
severe or critical (p < 0.001). The mean patient age was not
significantly different between the severe and critical groups. However,
IL-6, IL-8, procalcitonin (Table 2), CRP, ferroprotein (Figure 3A, 3B),
WBC count, and neutrophil count (Figure 4A, 4B) were all significantly
elevated in the critical group compared to severe. These data suggest
underlying differences in COVID-19 progression that is unrelated to age.
13.28.3 Significance
Given the inflammatory profile outlined in this study,
patients who have mild or severe COVID-19 pneumonia, who also have any
elevations in the inflammatory biomarkers listed above, should be
closely monitored for potential progression to critical status.
13.28.4 Credit
This review by JJF was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.29 An Effective CTL Peptide Vaccine for Ebola Zaire Based on Survivors’ CD8+ Targeting of a Particular Nucleocapsid Protein Epitope with Potential Implications for COVID-19 Vaccine Design
Vaccination of mice with a single dose of a 9-amino-acid peptide NP44-52
located in a conserved region of ebolavirus (EBOV) nucleocapsid protein
(NP) confers CD8+ T-cell-mediated immunity against mouse adapted EBOV
(maEBOV). Bioinformatic analyses predict multiple conserved CD8+ T cell
epitopes in the SARS-CoV-2 NP, suggesting that a similar approach may be
feasible for vaccine design against SARS-CoV-2.
The authors focus on a site within a 20-peptide region of EBOV NP which
was commonly targeted by CD8+ T cells in a group of EBOV survivors
carrying the HLA-A*30:01:01 allele. To justify the testing of specific
vaccine epitopes in a mouse challenge setting, the authors cite known
examples of human pathogen-derived peptide antigens that are also
recognized by C57BL/6 mice, as well as existing data surrounding known
mouse immunogenicity of peptides related to this EBOV NP region. Testing
3 distinct 9mer peptides over an 11 amino-acid window and comparing to
vaccination with the 11mer with a T-cell reactivity readout demonstrated
that optimizing peptide length and position for immunogenicity may be
crucial, likely due to suboptimal peptide processing and MHC-class-I
loading.
Vaccines for maEBOV challenge studies were constructed by packaging
NP44-52 in d,l poly(lactic-co-glycolic) acid microspheres. CpG was also
packaged within the microspheres, while Monophosphoryl Lipid A (a TLR4
ligand) was added to the injectate solution. A second peptide consisting
of a predicted MHC-II epitope from the EBOV VG19 protein was added using
a separate population of microspheres, and the formulation was injected
by intraperitoneal administration. The vaccine was protective against a
range of maEBOV doses up to at least 10,000 PFU. Survival was
anticorrelated with levels of IL6, MCP-1 (CCL2), IL9, and GM-CSF, which
recapitulated trends seen in human EBOV infection.
While HLA-A*30:01:01 is only present in a minority of humans, the
authors state that MHC binding algorithms predict NP44-52 to be a strong
binder of a set of more common HLA-A*02 alleles. The authors predict
that a peptide vaccine based on the proposed formulation could elicit
responses in up to 50% of people in Sudan or 30% of people in North
America.
SARS-CoV-2 NP, meanwhile, has conserved regions which may provide
peptide-vaccine candidates. Scanning the SARS-CoV-2 NP sequence for
HLA-binding 9mers identified 53 peptides with predicted binding affinity
< 500nM, including peptides that are predicted to bind to HLA-class-I
alleles of 97% of humans, 7 of which have previously been tested
in-vitro.
The results support previously appreciated correlations between certain
cytokines and disease severity, specifically IL6 which relates to
multiple trial therapies. Prediction of HLA-class-I binding of
SARS-CoV-2 NP peptides suggests the plausibility of a peptide vaccine
targeting conserved regions of SARS-CoV-2 NP although further validation
in previously infected patient samples will be essential.
13.29.3 Credit
Review by Andrew M. Leader as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.30 Epitope-based peptide vaccines predicted against novel coronavirus disease caused by SARS-CoV-2
This study employs a series of bioinformatic
pipelines to identify T and B cell epitopes on spike (S) protein of
SARS-CoV-2 and assess their properties for vaccine potential. To
identify B cell epitopes, they assessed structural accessibility,
hydrophilicity, and beta-turn and flexibility which are all factors that
promote their targeting by antibodies. To identify T cell epitopes, they
filtered for peptides with high antigenicity score and capacity to bind
3 or more MHC alleles. Using the protein digest server, they also
demonstrated that their identified T and B cell epitopes are stable,
having multiple non-digesting enzymes per epitope. Epitopes were also
determined to be non-allergenic and non-toxin as assessed by Allergen FP
1.0 and ToxinPred, respectively. For T cell epitopes, they assessed the
strength of epitope-HLA interaction via PepSite. Overall, they predict
four B cell and eleven T cell epitopes (two MHC I and nine MHC II
binding) to pass stringent computational thresholds as candidates for
vaccine development. Furthermore, they performed sequence alignment
between all identified SARS-CoV-2 S protein mutations and predicted
epitopes, and showed that the epitopes are conserved across 134 isolates
from 38 locations worldwide. However, they report that these conserved
epitopes may soon become obsolete given the known mutation rate of
related SARS-CoV is estimated to be 4x10-4/site/year, underscoring the
urgency of anti-viral vaccine development.
13.30.3 Limitations
While spike (S) protein may have a
critical role in viral entry into host cells and their epitope
prediction criterion were comprehensive, this study did not examine
other candidate SARS-CoV-2 proteins. This point is particularly
important given that a single epitope may not be sufficient to induce
robust immune memory, and recent approaches involve multi-epitope
vaccine design. Furthermore, their study only included a direct
implementation of various published methods, but did not validate
individual bioinformatic tools with controls to demonstrate robustness.
Finally, it is critical that these predicted epitopes are experimentally
validated before any conclusions can be drawn about their potential as
vaccine candidates or their clinical efficacy.
13.30.4 Significance
This study provides a computational framework to
rapidly identify epitopes that may serve as potential vaccine candidates
for treating SARS-CoV-2.
13.30.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.31 The definition and risks of Cytokine Release Syndrome-Like in 11 COVID-19-Infected Pneumonia critically ill patients: Disease Characteristics and Retrospective Analysis
This study describes the occurrence of a cytokine release syndrome-like
(CRSL) toxicity in ICU patients with COVID-19 pneumonia. The median time
from first symptom to acute respiratory distress syndrome (ARDS) was 10
days. All patients had decreased CD3, CD4 and CD8 cells, and a
significant increase of serum IL-6. Furthermore, 91% had decreased NK
cells. The changes in IL-6 levels preceded those in CD4 and CD8 cell
counts. All of these parameters correlated with the area of pulmonary
inflammation in CT scan images. Mechanical ventilation increased the
numbers of CD4 and CD8 cells, while decreasing the levels of IL-6, and
improving the immunological parameters.
13.31.3 Limitations
The number of patients included in this retrospective single center
study is small (n=11), and the follow-up period very short (25 days).
Eight of the eleven patients were described as having CRSL, and were
treated by intubation (7) or ECMO (2). Nine patients were still in the
intensive care unit at the time of publication of this article, so their
disease outcome is unknown.
13.31.4 Significance
The authors define a cytokine release syndrome-like toxicity in patients
with COVID-19 with clinical radiological and immunological criteria: 1)
decrease of circulating CD4, CD8 and NK cells; 2) substantial increase
of IL-6 in peripheral blood; 3) continuous fever; 4) organ and tissue
damage. This event seems to occur very often in critically ill patients
with COVID-19 pneumonia. Interestingly, the increase of IL-6 in the
peripheral blood preceded other laboratory alterations, thus, IL-6 might
be an early biomarker for the severity of COVID-19 pneumonia. The
manuscript will require considerable editing for organization and
clarity.
13.31.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.32 Clinical characteristics of 36 non-survivors with COVID-19 in Wuhan, China
This is a simple study reporting clinical
characteristics of patients who did not survive COVID-19. All patients
(mean age=69.22 years) had acute respiratory distress syndrome (ARDS)
and their median time from onset to ARDS was 11 days. The median time
from onset to death was 17 days. Most patients were older male (70%
male) with co-morbidities and only 11 % were smokers. 75% patients
showed bilateral pneumonia. Many patients had chronic diseases,
including hypertension (58.33%). cardiovascular disease (22.22%) and
diabetes (19.44%). Typical clinical feature measured in these patients
includes lymphopenia and elevated markers of inflammation.
13.32.3 Limitations
As noted by the authors, the conclusions of this study are
very limited because this is single-centered study focusing on a small
cohort of patients who did not survive. Many clinical parameters
observed by the authors (such* as increase levels of serum CRP, PCT,
IL-6) have also been described in other COVID19 patients who survived
the infection
13.32.4 Significance
This study is essentially descriptive and may be useful for
clinical teams monitoring COVID19 patients.
13.32.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.33 Risk Factors Related to Hepatic Injury in Patients with Corona Virus Disease 2019
Based on a retrospective study of 85 hospitalized COVID
patients in a Beijing hospital, authors showed that patients with
elevated ALT levels (n = 33) were characterized by significantly higher
levels of lactic acid and CRP as well as lymphopenia and hypoalbuminemia
compared to their counterparts with normal ALT levels. Proportion of
severe and critical patients in the ALT elevation group was
significantly higher than that of normal ALT group. Multivariate
logistic regression performed on clinical factors related to ALT
elevation showed that CRP \(\geq\) 20mg/L and low lymphocyte count
(<1.1*10^9 cells/L) were independently related to ALT elevation—a
finding that led the authors to suggest cytokine storm as a major
mechanism of liver damage.
13.33.3 Limitations
The article’s most attractive claim that liver damage seen in COVID
patients is caused by cytokine storm (rather than direct infection of
the liver) hinges solely on their multivariate regression analysis.
Without further mechanistic studies a) demonstrating how high levels of
inflammatory cytokines can induce liver damage and b) contrasting types
of liver damage incurred by direct infection of the liver vs.
system-wide elevation of inflammatory cytokines, their claim remains
thin. It is also worth noting that six of their elevated ALT group
(n=33) had a history of liver disease (i.e. HBV infection, alcoholic
liver disease, fatty liver) which can confound their effort to pin down
the cause of hepatic injury to COVID.
13.33.4 Significance
Limited. This article confirms a rich body
of literature describing liver damage and lymphopenia in COVID patients.
13.33.5 Credit
Review by Chang Moon as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.34 Detectable serum SARS-CoV-2 viral load (RNAaemia) is closely associated with drastically elevated interleukin 6 (IL-6) level in critically ill COVID-19 patients
48 adult patients diagnosed with Covid19 according to Chinese guidelines
for Covid19 diagnosis and treatment version 6 were included in this
study. Patients were further sub-divided into three groups based on
clinical symptoms and disease severity: (1) mild, positive Covid19 qPCR
with no or mild clinical symptoms (fever; respiratory; radiological
abnormalities); (2) severe, at least one of the following: shortness of
breath/respiratory rate >30/min, oxygen saturation SaO2<93%,
Horowitz index paO2/FiO2 < 300 mmHg (indicating moderate pulmonary
damage); and (3) critically ill, at least one additional complicating
factor: respiratory failure with need for mechanical ventilation;
systemic shock; multi-organ failure and transfer to ICU. Serum samples
and throat-swaps were collected from all 48 patients enrolled.
SARS-CoV-2 RNA was assessed by qPCR with positive results being defined
as Ct values < 40, and serum interleukin-6 (IL-6) was quantified
using a commercially available detection kit. Briefly, patient
characteristics in this study confirm previous reports suggesting that
higher age and comorbidities are significant risk factors of clinical
severity. Of note, 5 out of 48 of patients (10.41%), all in the
critically ill category, were found to have detectable serum SARS-CoV-2
RNA levels, so-called RNAaemia. Moreover, serum IL-6 levels in these
patients were found to be substantially higher and this correlated with
the presence of detectable SARS-CoV-2 RNA levels. The authors
hypothesize that viral RNA might be released from acutely damages
tissues in moribund patients during the course of Covid19 and that
RNaemia along with IL-6 could potentially be used as a prognostic
marker.
13.34.3 Limitations
While this group’s report generally confirms some of the major findings
of a more extensive study, published in early February 2020, [1281],
there are limitations that should be taken into account. First, the
number of patients enrolled is relatively small; second, interpretation
of these data would benefit from inclusion of information about study
specifics as well as providing relevant data on the clinical course of
these patients other than the fact that some were admitted to ICU (i.e.
demographics on how many patients needed respiratory support, dialysis,
APACHE Ii/III or other standard ICU scores as robust prognostic markers
for mortality etc). It also remains unclear at which time point the
serum samples were taken, i.e. whether at admission, when the diagnosis
was made or during the course of the hospital stay (and potentially
after onset of therapy, which could have affected both IL-6 and RNA
levels). The methods section lacks important information on the qPCR
protocol employed, including primers and cycling conditions used. From a
technical point of view, Ct values >35 seem somewhat non-specific
(although Ct <40 was defined as the CDC cutoff as well) indicating
that serum RNA levels are probably very low, therefore stressing the
need for highly specific primers and high qPCR efficiency. In addition,
the statistical tests used (t-tests, according to the methods section)
do not seem appropriate as the organ-specific data such as BUN and
troponin T values seem to be not normally distributed across groups (n=
5 RNAaemia+ vs. n= 43 RNAaemia-). Given the range of standard deviations
and the differences in patient sample size, it is difficult to believe
that these data are statistically significantly different.
13.34.4 Significance
This study is very rudimentary and lacks a lot of relevant clinical
details. However, it corroborates some previously published observations
regarding RNAemia and IL-6 by another group. Generally, regarding future
studies, it would be important to address the question of IL-6 and other
inflammatory cytokine dynamics in relation to Covid19 disease kinetics
(high levels of IL-6, IL-8 and plasma leukotriene were shown to have
prognostic value at the onset of ARDS ; serum IL-2 and IL-15 have been
associated with mortality; reviewed by Chen W & Ware L, Clin Transl Med.
2015 [1292]).
13.34.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.35 Lymphopenia predicts disease severity of COVID-19: a descriptive and predictive study
Based on a retrospective study of 162 COVID patients from a
local hospital in Wuhan, China, the authors show an inverse correlation
between lymphocyte % (LYM%) of patients and their disease severity. The
authors have also tracked LYM% of 70 cases (15 deaths; 15 severe; 40
moderate) throughout the disease progression with fatal cases showing no
recovery of lymphocytes ( <5%) even after 17-19 days post-onset. The
temporal data of LYM % in COVID patients was used to construct a
Time-Lymphocyte% model which is used to categorize and predict patients’
disease severity and progression. The model was validated using 92
hospitalized cases and kappa statistic test was used to assess agreement
between predicted disease severity and the assigned clinical severity (k
= 0.49).
13.35.3 Limitations
Time-Lymphocyte % Model (TLM) that authors have proposed as
a predictive model for clinical severity is very simple in its
construction and derives from correlative data of 162 patients. In order
for the model to be of use, it needs validation using a far more robust
data set and possibly a mechanistic study on how COVID leads to
lymphopenia in the first place. In addition, it should be noted that no
statistical test assessing significance of LYM % values between disease
severities was performed.
13.35.4 Significance
This article is of limited significance as
it simply reports similar descriptions of COVID patients made in
previous literature that severe cases are characterized by lymphopenia.
13.35.5 Credit
Review by Chang Moon as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.36 The potential role of IL-6 in monitoring severe case of coronavirus disease 2019
Study on blood biomarkers on 80 COVID19 patients (69 severe
and 11 non-severe). Patients with severe symptoms at admission
(baseline) showed obvious lymphocytopenia and significantly increased
interleukin-6 (IL-6) and CRP, which was positively correlated with
symptoms severity. IL-6 at baseline positively correlates with CRP, LDH,
ferritin and D-Dimer abundance in blood.
Longitudinal analysis of 30 patients (before and after treatment) showed
significant reduction of IL-6 in remission cases.
13.36.3 Limitations
Limited sample size at baseline, especially for the
non-severe leads to question on representativeness. The longitudinal
study method is not described in detail and suffers from
non-standardized treatment. Limited panel of pro-inflammatory cytokine
was analyzed. Patients with severe disease show a wide range of altered
blood composition and biomarkers of inflammation, as well as differences
in disease course (53.6% were cured, about 10% developed acute
respiratory distress syndrome). The authors comment on associations
between IL-6 levels and outcomes, but these were not statistically
significant (maybe due to the number of patients, non-standardized
treatments, etc.) and data is not shown. Prognostic biomarkers could
have been better explored. Study lacks multivariate analysis.
13.36.4 Significance
IL-6 could be used as a pharmacodynamic
marker of disease severity. Cytokine Release Syndrome (CRS) is a
well-known side effect for CAR-T cancer therapy and there are several
effective drugs to manage CRS. Drugs used to manage CRS could be tested
to treat the most severe cases of COVID19.
13.36.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.37 Clinical and Laboratory Profiles of 75 Hospitalized Patients with Novel Coronavirus Disease 2019 in Hefei, China
The authors of this study provide a comprehensive analysis of clinical
laboratory assessments in 75 patients (median age 47 year old)
hospitalized for Corona virus infection in China measuring differential
blood counts including T-cell subsets (CD4, CD8), coagulation function,
basic blood chemistry, of infection-related biomarkers including CRP,
Procalcitonin (PCT) (Precursor of calcitonin that increases during
bacterial infection or tissue injury), IL-6 and erythrocyte
sedimentation rate as well as clinical parameters. Among the most common
hematological changes they found increased neutrophils, reduced CD4 and
CD8 lymphocytes, increased LDH, CRP and PCT
When looking at patients with elevated IL-6, the authors describe
significantly reduced CD4 and CD8 lymphocyte counts and elevated CRP and
PCT levels were significantly increased in infected patients suggesting
that increased IL-6 may correlate well with disease severity in COVID-19
infections
13.37.3 Limitations
The authors performed an early assessment of clinical standard
parameters in patients infected with COVID-19. Overall, the number of
cases (75) is rather low and the snapshot approach does not inform about
dynamics and thus potential relevance in the assessment of treatment
options in this group of patients.
13.37.4 Significance
The article summarizes provides a good summary of some of the common
changes in immune cells inflammatory cytokines in patients with a
COVID-19 infection and. Understanding how these changes can help predict
severity of disease and guide therapy including IL-6 cytokine receptor
blockade using Tocilizumab or Sarilumab will be important to explore.
13.37.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.38 Exuberant elevation of IP-10, MCP-3 and IL-1ra during SARS-CoV-2 infection is associated with disease severity and fatal outcome
Plasma cytokine analysis (48 cytokines) was
performed on COVID-19 patient plasma samples, who were sub-stratified as
severe (N=34), moderate (N=19), and compared to healthy controls (N=8).
Patients were monitored for up to 24 days after illness onset: viral
load (qRT-PCR), cytokine (multiplex on subset of patients), lab tests,
and epidemiological/clinical characteristics of patients were reported.
13.38.3 Main Findings
Many elevated cytokines with COVID-19 onset compared to healthy
controls (IFNy, IL-1Ra, IL-2Ra, IL-6, IL-10, IL-18, HGF, MCP-3, MIG,
M-CSF, G-CSF, MIG-1a, and IP-10).
IP-10, IL-1Ra, and MCP-3 (esp. together) were associated with
disease severity and fatal outcome.
IP-10 was correlated to patient viral load (r=0.3006, p=0.0075).
IP-10, IL-1Ra, and MCP-3 were correlated to loss of lung function
(PaO2/FaO2 (arterial/atmospheric O2) and Murray Score (lung
injury) with MCP-3 being the most correlated (r=0.4104 p<0.0001
and r=0.5107 p<0.0001 respectively).
Viral load (Lower Ct Value from qRT-PCR) was associated with
upregulated IP-10 only (not IL-1Ra or MCP-3) and was mildly
correlated with decreased lung function: PaO2/FaO2
(arterial/atmospheric O2) and Murray Score (lung injury).
Lymphopenia (decreased CD4 and CD8 T cells) and increased neutrophil
correlated w/ severe patients.
Complications were associated with COVID severity (ARDS, hepatic
insufficiency, renal insufficiency).
13.38.4 Limitations
Collection time of clinical data and lab results
not reported directly (likely 4 days (2,6) after illness onset), making
it very difficult to determine if cytokines were predictive of patient
outcome or reflective of patient compensatory immune response (likely
the latter). Small N for cytokine analysis (N=2 fatal and N=5
severe/critical, and N=7 moderate or discharged). Viral treatment
strategy not clearly outlined.
13.38.5 Expanded Results
NOTE: Moderate COVID-19 was classified by fever, respiratory
manifestations, and radiological findings consistent with pneumonia
while severe patients had one or more of the following: 1) respiratory
distraction, resting O2 saturation, or 3) arterial PaO2/FiO2 < 300
mmHg.
Significant elevation of cytokines observed in COVID patients
compared to healthy controls: IFNy, IL-1Ra, IL-2Ra, IL-6, IL-10,
IL-18, HGF, MCP-3, MIG, M-CSF, G-CSF, MIG-1a, and IP-10.
Severity was correlated with increase in measured IP-10, MCP-3,
and IL-Ra as measure by area under the curve analysis during
sample timecourse (2-24 days after illness onset).
IL-1Ra incr. significant 0-7 days after onset, MCP-3 signif.
upregulated throughout observation timecourse, and IP-10 increased
and upregulated throughout (trending downwards over time).
The three cytokines together (IP-10, IL-1Ra, and MCP-3 AUC) served
as the best predictors of disease deterioration and fatal outcome.
No significant differences between moderate/severe observed between
groups in IL-2Ra, IL-6, IL-10, IL-18, CTACK, G-CSF, HGF, M-CSF,
MIP-1a, MIG, and IFNy at any timepoints.
Viral load (Lower Ct Value from qRT-PCR) was associated with
upregulated IP-10 only (not IL-1Ra or MCP-3) and was highly
correlated with decreased lung function: PaO2/FaO2
(arterial/atmospheric O2) and Murray Score (lung injury).
Antibodies against these cytokines (esp. anti-IP-10) may serve as
a potential treatment for amelioration of COVID-19 (and associated
ARDS).
Lab results:
Decreased lymphocytes (%) in all patients – lymphopenia corr. w/
severe patients
Decreased CD4 and CD8 T cells – no monocyte or
eosino/basophil % measured
Increased neutrophils (%)
Increased BUN (mmol/L) – other kidney markers, liver markers, and
LDH were not significantly different between groups and were not
compared to healthy controls.
Clinical features (between moderate vs. severe patient groups):
Complications were associated with severity (ARDS, hepatic
insufficiency, renal insufficiency).
Coexisting conditions between groups were not significantly
different (chronic heart/lung/renal/liver disease, diabetes, or
cancer) and patient time courses (onset to admission and onset to
viral tx) also not significantly different – 4 days (2, 6) on
average for admission and 4 (3,7) for antiviral.
Increased corticosteroids and mechanical/ invasive mechanical
ventilation in severe patients.
Increased median age in severe group (Median (Range = 63.5 (42-74)
vs. 51 (22-78)) and patients > 60 yrs had higher ratio of
severe patients as compared patients 16-59 yrs.
Higher incidence of fever is severe patients (91.2 vs. 68.4%),
myalgia (57.7 vs. 48.1%), and chill (17.6% vs. 0%).
No differences in cough, headache, nausea/vomiting, or diarrhea.
13.38.6 Significance
Outline of pathological time course (implicating innate
immunity esp.) and identification key cytokines associated with disease
severity and prognosis (+ comorbidities). Anti-IP-10 as a possible
therapeutic intervention (ex: Eldelumab).
13.38.7 Credit
Review by Natalie Vaninov as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.39 Antibody responses to SARS-CoV-2 in patients of novel coronavirus disease 2019
This study examined antibody responses in
the blood of COVID-19 patients during the early SARS CoV2 outbreak in
China. Total 535 plasma samples were collected from 173 patients (51.4%
female) and were tested for seroconversion rate using ELISA. Authors
also compared the sensitivity of RNA and antibody tests over the course
of the disease . The key findings are:
Among 173 patients, the seroconversion rates for total antibody
(Ab), IgM and IgG were 93.1% (161/173), 82.7% (143/173) and 64.7%
(112/173), respectively.
The seroconversion sequentially appeared for Ab, IgM and then IgG,
with a median time of 11, 12 and 14 days, respectively. Overall, the
seroconversion of Ab was significantly quicker than that of IgM (p =
0.012) and IgG (p < 0.001). Comparisons of seroconversion rates
between critical and non-critical patients did not reveal any
significant differences.
RNA tests had higher sensitivity in early phase and within 7 days of
disease onset than antibody assays (66.7% Vs 38.3% respectively).
The sensitivity of the Ab assays was higher 8 days after disease
onset, reached 90% at day 13 and 100% at later time points (15-39
days). In contrast, RNA was only detectable in 45.5% of samples at
days 15-39.
In patients with undetectable RNA in nasal samples collected during
day 1-3, day 4-7, day 8-14 and day 15-39 since disease onset, 28.6%
(2/7), 53.6% (15/28), 98.2% (56/57) and 100% (30/30) had detectable
total Ab titers respectively Combining RNA and antibody tests
significantly raised the sensitivity for detecting COVID-19 patients
in different stages of the disease (p < 0.001).
There was a strong positive correlation between clinical severity
and antibody titer 2-weeks after illness onset.
Dynamic profiling of viral RNA and antibodies in representative
COVID-19 patients (n=9) since onset of disease revealed that
antibodies may not be sufficient to clear the virus. It should be
noted that increases in of antibody titers were not always
accompanied by RNA clearance.
13.39.3 Limitations
Because different types of ELISA assays were used for
determining antibody concentrations at different time points after
disease onset, sequential seroconversion of total Ab, IgM and IgG may
not represent actual temporal differences but rather differences in the
affinities of the assays used. Also, due to the lack of blood samples
collected from patients in the later stage of illness, how long the
antibodies could last remain unknown. For investigative dynamics of
antibodies, more samples were required.
13.39.4 Significance
Total and IgG antibody titers could be used to understand
the epidemiology of SARS CoV-2 infection and to assist in determining
the level of humoral immune response in patients.
The findings provide strong clinical evidence for routine serological
and RNA testing in the diagnosis and clinical management of COVID-19
patients. The understanding of antibody responses and their half-life
during and after SARS CoV2 infection is important and warrants further
investigations.
13.39.5 Credit
This review was undertaken by Zafar Mahmood and edited by K Alexandropoulos as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.40 Restoration of leukomonocyte counts is associated with viral clearance in COVID-19 hospitalized patients
The authors collected data on 25 COVID-19 patients (n=11 men, n=14
women) using standard laboratory tests and flow cytometry. All patients
were treated with antibiotics. Twenty-four of the 25 patients were also
treated with anti-viral Umefinovir and 14 of the patients were treated
with corticosteroids. 14 patients became negative for the virus after
8-14 days of treatment. The same treatment course was extended to 15-23
days for patients who were still positive for the virus at day 14.
The authors found a negative association between age and resolution of
infection. Patients with hypertension, diabetes, malignancy or chronic
liver disease were all unable to clear the virus at day 14, though not
statistically significant.
Elevated procalcitonin and a trend for increased IL-6 were also found in
peripheral blood prior to the treatment.
A trend for lower NK cell, T cell and B cell counts in patients was also
reported. B cell, CD4 and CD8 T cell counts were only increased upon
treatment in patients who cleared the virus. NK cell frequencies
remained unchanged after treatment in all the patients.
13.40.3 Limitations
73% of the patients who remained positive for SARS-CoV2 after the 1st
treatment, and 43% of all patients who cleared the virus were treated
with corticosteroids. Corticosteroids have strong effects on the immune
compartment in blood [1299]. The authors should have accounted for
corticosteroid treatment when considering changes in T, NK and B cell
frequencies.
Assessing if IL-6 concentrations were back to baseline levels following
treatment would have provided insights into the COVID-19 cytokine storm
biology. Patients with higher baseline levels of IL-6 have been reported
to have lower CD8 and CD4 T cell frequencies [1295]. Correlating IL-6 with
cell counts before and after treatment would thus have also been of
interest. The report of the laboratory measures in table 2 is incomplete
and should include the frequencies of patients with increased/decreased
levels for each parameter.
Correction is needed for the 1st paragraph of the discussion as data
does not support NK cell restoration upon treatment in patients who
cleared the virus. NK cells remain unchanged after the 1st treatment
course and only seem to increase in 2 out of 6 donors after the 2nd
treatment course in those patients.
13.40.4 Significance
Previous reports suggest an association between disease severity and
elevated IL-6 or pro-calcitonin concentrations in COVID-19
patients [1291,1300]. IL-6 receptor blockade is also being administered to
patients enrolled in clinical trials (NCT04317092). This report thus
contributes to highlight elevated concentrations of these analytes in
COVID-19 patients. Mechanisms underlying the association between viral
clearance and restoration of the T cell and B cell frequencies suggests
viral-driven immune dysregulation, which needs to be investigated in
further studies.
13.40.5 Credit
Review by Bérengère Salomé as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.41 Clinical findings in critically ill patients infected with SARS-CoV-2 in Guangdong Province, China: a multi-center, retrospective, observational study
This work analyses laboratory and clinical data from 45 patients treated
in the in ICU in a single province in China. Overall, 44% of the
patients were intubated within 3 days of ICU admission with only 1
death.
Lymphopenia was noted in 91% of patient with an inverse correlation with
LDH.
Lymphocyte levels are negatively correlated with Sequential Organ
Failure Assessment (SOFA) score (clinical score, the higher the more
critical state), LDH levels are positively correlated to SOFA score.
Overall, older patients (>60yo), with high SOFA score, high LDH
levels and low lymphocytes levels at ICU admission are at higher risk of
intubation.
Of note, convalescent plasma was administered to 6 patients but due to
limited sample size no conclusion can be made.
13.41.3 Limitations
While the study offers important insights into disease course and
clinical lab correlates of outcome, the cohort is relatively small and
is likely skewed towards a less-severe population compared to other ICU
reports given the outcomes observed. Analysis of laboratory values and
predictors of outcomes in larger cohorts will be important to make
triage and treatment decisions. As with many retrospective analyses,
pre-infection data is limited and thus it is not possible to understand
whether lymphopenia was secondary to underlying comorbidities or
infection.
Well-designed studies are necessary to evaluate the effect of
convalescent plasma administration.
13.41.4 Significance
This clinical data enables the identification of at-risk
patients and gives guidance for research for treatment options. Indeed,
further work is needed to better understand the causes of the
lymphopenia and its correlation with outcome.
13.41.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.42 Immune Multi-epitope vaccine design using an immunoinformatics approach for 2019 novel coronavirus in China (SARS-CoV-2)
Using in silico bioinformatic tools, this study identified putative
antigenic B-cell epitopes and HLA restricted T-cell epitopes from the
spike, envelope and membrane proteins of SARS-CoV-2, based on the genome
sequence available on the NCBI database. T cell epitopes were selected
based on predicted affinity for the more common HLA-I alleles in the
Chinese population. Subsequently, the authors designed vaccine peptides
by bridging selected B-cell epitopes and adjacent T-cell epitopes.
Vaccine peptides containing only T-cell epitopes were also generated.
From 61 predicted B-cell epitopes, only 19 were exposed on the surface
of the virion and had a high antigenicity score. A total of 499 T-cell
epitopes were predicted. Based on the 19 B-cell epitopes and their 121
adjacent T-cell epitopes, 17 candidate vaccine peptides were designed.
Additionally, another 102 vaccine peptides containing T-cell epitopes
only were generated. Based on the epitope counts and HLA score, 13 of
those were selected. Thus, a total of 30 peptide vaccine candidates were
designed.
13.42.3 Limitations
While this study provides candidates for the development of vaccines
against SARS-CoV-2, in vitro and in vivo trials are required to validate
the immunogenicity of the selected B and T cell epitopes. This could be
done using serum and cells from CoV-2-exposed individuals, and in
preclinical studies. The implication of this study for the current
epidemic are thus limited. Nevertheless, further research on this field
is greatly needed.
13.42.4 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.43 Clinical Features of Patients Infected with the 2019 Novel Coronavirus (COVID-19) in Shanghai, China
This single-center cohort study analyzes the clinical and
laboratory features of 198 patients with confirmed COVID-19 infection in
Shanghai, China and correlated these parameters with clinical disease
severity, including subsequent intensive care unit (ICU) admission. 19
cases (9.5%) required ICU admission after developing respiratory failure
or organ dysfunction. Age, male sex, underlying cardiovascular disease,
and high symptom severity (high fever, dyspnea) were all significantly
correlated with ICU admission. Additionally, ICU admission was more
common in patients who presented with lymphopenia and elevated
neutrophil counts, among other laboratory abnormalities. Flow cytometric
analysis revealed that patients admitted to the ICU had significantly
reduced circulating CD3+ T cell, CD4+ T cell, CD8+ T cell, and CD45+
leukocyte populations compared to the cohort of patients not requiring
ICU admission.
13.43.3 Limitations
The limitations of this study include the relatively
small sample size and lack of longitudinal testing. The authors also did
not assess whether respiratory comorbidity – such as asthma or chronic
obstructive lung disease – in addition to immunosuppression affected ICU
admission likelihood.
13.43.4 Significance
COVID-19 has already sickened thousands across the globe,
though the severity of these infections is markedly diverse, ranging
from mild symptoms to respiratory failure requiring maximal
intervention. Understanding what clinical, laboratory, and immunologic
factors predict the clinical course of COVID-19 infection permits
frontline providers to distribute limited medical resources more
effectively.
13.43.5 Credit
Review by Andrew Charap as part of a project by students, postdocs and
faculty at the Immunology Institute of the Icahn School of Medicine at
Mount Sinai.
13.44 Serological detection of 2019-nCoV respond to the epidemic: A useful complement to nucleic acid testing
This study showed that both anti-2019-nCov IgM and IgG were detected by
automated chemiluminescent immunoassay in the patients who had been
already confirmed as positive by nucleic acid detection, while single
positivity of IgM or IgG were detected in a very few cases in the other
population including 225 non-COVID-19 cases. In addition to the increase
of anti-2019-nCov IgM 7-12 days after morbidity, the increase of IgG was
detected in three patients with COVID-19 within a very short of time
(0-1 day).
13.44.3 Limitations
The limitation of this study is only
3 confirmed COVID-19 cases were included, so that the relationship
between anti-2019-nCov antibodies and disease progression might not be
clearly defined. Another limitation is that they did not show the course
of 2019-nCov specific antibodies in the cases with positive for COVID-19
but without clinical symptoms.
13.44.4 Significance
The detection of anti-2019-nCov antibodies can be an alternative method
to diagnose and treat COVID-19 more comprehensively by distinguish non
COVID-19 patients. It may be helpful to understand the course of
individual cases with COVID-19 to predict the prognosis if more cases
will be evaluated.
13.44.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.45 Human Kidney is a Target for Novel Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Infection
Analyzing the eGFR (effective glomerular flow rate) of 85 Covid-19
patients and characterizing tissue damage and viral presence in
post-mortem kidney samples from 6 Covid-19 patients, the authors
conclude that significant damage occurs to the kidney, following
Covid-19 infection. This is in contrast to the SARS infection from the
2003 outbreak. They determine this damage to be more prevalent in
patients older than 60 years old, as determined by analysis of eGFR. H&E
and IHC analysis in 6 Covid-19 patients revealed that damage was in the
tubules, not the glomeruli of the kidneys and suggested that macrophage
accumulation and C5b-9 deposition are key to this process.
13.45.3 Limitations
Severe limitations include that the H&E and IHC samples were performed
on post-mortem samples of unknown age, thus we cannot assess how/if age
correlates with kidney damage, upon Covid-19 infection. Additionally,
eGFR was the only in-vivo measurement. Blood urea nitrogen and
proteinuria are amongst other measurements that could have been obtained
from patient records. An immune panel of the blood was not performed to
assess immune system activation. Additionally, patients are only from
one hospital.
13.45.4 Significance
This report makes clear that kidney damage is prevalent
in Covid-19 patients and should be accounted for.
13.45.5 Credit
Review by Dan Fu Ruan, Evan Cody and Venu Pothula as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine at Mount Sinai.
13.46 COVID-19 early warning score: a multi-parameter screening tool to identify highly suspected patients
The aim of this study was to identify diagnostic or prognostic criteria
which could identify patients with COVID-19 and predict patients who
would go on to develop severe respiratory disease. The authors use EMR
data from individuals taking a COVID-19 test at Zhejiang hospital, China
in late January/Early February. A large number of clinical parameters
were different between individuals with COVID-19 and also between
‘severe’ and ‘non-severe’ infections and the authors combine these into
a multivariate linear model to derive a weighted score, presumably
intended for clinical use.
13.46.3 Limitations
Unfortunately, the paper is lacking a lot of crucial information, making
it impossible to determine the importance or relevance of the findings.
Most importantly, the timings of the clinical measurements are not
described relative to the disease course, so it is unclear if the
differences between ‘severe’ and ‘non-severe’ infections are occurring
before progression to severe disease (which would make them useful
prognostic markers), or after (which would not).
13.46.4 Significance
This paper is one of many retrospective studies coming from hospitals in
China studying individuals with COVID-19. Because of the sparse
description of the study design, this paper offers little new
information. However, studies like this could be very valuable and we
would strongly encourage the authors to revise this manuscript to
include more information about the timeline of clinical measurements in
relation to disease onset and more details of patient outcomes.
13.46.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.47 LY6E impairs coronavirus fusion and confers immune control of viral disease
Screening a cDNA library of >350 human interferon-stimulated genes
for antiviral activity against endemic human coronavirus HCoV-229E
(associated with the common cold), Pfaender S & Mar K et al. identify
lymphocyte antigen 6 complex, locus E (Ly6E) as an inhibitor of cellular
infection of Huh7 cells, a human hepatoma cell line susceptible to
HCoV-229E and other coronaviruses. In a series of consecutive in vitro
experiments including both stable Ly6E overexpression and
CRISPR-Cas9-mediated knockout the authors further demonstrate that Ly6E
reduces cellular infection by various other coronaviruses including
human SARS-CoV and SARS-CoV-2 as well as murine CoV mouse hepatitis
virus (MHV). Their experiments suggest that this effect is dependent on
Ly6E inhibition of CoV strain-specific spike protein-mediated membrane
fusion required for viral cell entry.
To address the function of Ly6E in vivo, hematopoietic stem
cell-specific Ly6E knock-out mice were generated by breeding Ly6Efl/fl
mice (referred to as functional wild-type mice) with transgenic
Vav-iCre mice (offspring referred to as Ly6E HSC ko mice); wild-type
and Ly6E HSC ko mice of both sexes were infected intraperitoneally with
varying doses of the natural murine coronavirus MHV, generally causing a
wide range of diseases in mice including hepatitis, enteritis and
encephalomyelitis. Briefly, compared to wild-type controls, mice lacking
hematopoietic cell-expressed Ly6E were found to present with a more
severe disease phenotype as based on serum ALT levels (prognostic of
liver damage), liver histopathology, and viral titers in the spleen.
Moreover, bulk RNAseq analysis of infected liver and spleen tissues
indicated changes in gene expression pathways related to tissue damage
and antiviral immune responses as well as a reduction of genes
associated with type I IFN response and inflammation. Finally, the
authors report substantial differences in the numbers of hepatic and
splenic APC subsets between wild-type and knockout mice following MHV
infection and show that Ly6E-deficient B cells and to a lesser extent
also DCs are particularly susceptible to MHV infection in vitro.
13.47.3 Limitations
Experiments and data in this study are presented in an overall logical
and coherent fashion; however, some observations and the conclusions
drawn are problematic and should be further addressed & discussed by the
authors. Methodological & formal limitations include relatively low
replicate numbers as well as missing technical replicates for some in
vitro experiments (cf. Fig. legend 1; Fig. legend 2e); the omission
of “outliers” in Fig. legend 2 without an apparent rationale as to why
this approach was chosen; the lack of detection of actual Ly6E protein
levels in Ly6E HSC ko or wild-type mice; and most importantly, missing
information on RNAseq data collection & analysis in the method section
and throughout the paper. A more relevant concern though is that the
interpretation of the experimental data presented and the language used
tend to overrate and at times overgeneralize findings: for example,
while the authors demonstrate statistically significant, Ly6E-mediated
reduction of coronavirus titers in stable cells lines in vitro, it
remains unclear whether a viral titer reduction by one log decade would
be of actual biological relevance in face of high viral titers in
vivo. After high-dose intraperitoneal MHV infection in vivo, early
viral titers in Ly6E HSC knockout vs. wt mice only showed an elevation
in the spleen (~1.5 log decades) but not liver of the ko mice (other
tissue not evaluated), and while ko mice presented with only modestly
increased liver pathology, both male and female ko mice exhibited
significantly higher mortality. Thus, the manuscript tile statement that
“Ly6E … confers immune control of viral disease” is supported by only
limited in vivo data, and gain-of-function experiments (eg. Ly6E
overexpression) were not performed. Of additional note here, tissue
tropism and virulence differ greatly among various MHV strains and
isolates whereas dose, route of infection, age, genetic background and
sex of the mice used may additionally affect disease outcome and
phenotype (cf. Taguchi F & Hirai-Yuki A,
https://doi.org/10.3389/fmicb.2012.00068; Kanolkhar A et al,
https://jvi.asm.org/content/
83/18/9258). Observations
attributed to hematopoietic stem cell-specific Ly6E deletion could
therefore be influenced by the different genetic backgrounds of floxed
and cre mice used, and although it appears that littermates wt and ko
littermates were used in the experiments, the potentially decisive
impact of strain differences should at least have been discussed. Along
these lines, it should also be taken into account that the majority of
human coronaviruses cause respiratory symptoms, which follow a different
clinical course engaging other primary cellular mediators than the
hepatotropic murine MHV disease studied here. It therefore remains
highly speculative how the findings reported in this study will
translate to human disease and it would therefore be important to test
other routes of MHV infection and doses that have been described to
produce a more comparable phenotype to human coronavirus disease (cf.
Kanolkhar A et al, https://jvi.asm.org/content/
83/18/9258). Another
important shortcoming of this study is the lack of any information on
functional deficits or changes in Ly6E-deficient immune cells and how
this might relate to the phenotype observed. Overall, the in vitro
experiments are more convincing than the in vivo studies which appear
somewhat limited.
13.47.4 Significance
Despite some shortcomings, the experiments performed in this study
suggest a novel and somewhat unexpected role of Ly6E in the protection
against coronaviruses across species. These findings are of relevance
and should be further explored in ongoing research on potential
coronavirus therapies. Yet an important caveat pertains to the authors’
suggestion that “therapeutic mimicking of Ly6E action” may constitute a
first line of defense against novel coronaviruses since their own prior
work demonstrated that Ly6E can enhance rather than curtail infection
with influenza A and other viruses.
13.47.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.48 A preliminary study on serological assay for severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) in 238 admitted hospital patients
While RT-PCR is being used currently to routinely diagnose infection
with SARS-CoV-2, there are significant limitations to the use of a
nucleic acid test that lead to a high false-negative rate. This
article describes ELISAs that can measure IgM and IgG antibodies
against the N protein of SARS-CoV-2 to test samples from 238
patients (153 positive by RT-PCR and 85 negative by RT-PCR) at
different times after symptom onset. The positivity rate of the IgM
and/or IgG ELISAs was greater than that of the RT-PCR (81.5%
compared to 64.3%) with similar positive rates in the confirmed and
suspected cases (83% and 78.8%, respectively), suggesting that many
of the suspected but RT-PCR-negative cases were also infected. The
authors also found that the ELISAs have higher positive rates later
after symptom onset while RT-PCR is more effective as a diagnostic
test early during the infection.
13.48.3 Limitations
I cannot identify any limitations to this study.
13.48.4 Significance
The authors make a strong case for using a combination of ELISA and
RT-PCR for diagnosis of infection with SARS-CoV-2, especially
considering the dynamics of positivity rates of RT-PCR and ELISA.
Fewer false-negative diagnoses would improve infection control and
patient management.
13.48.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.49 Monoclonal antibodies for the S2 subunit of spike of SARS-CoV cross-react with the newly-emerged SARS-CoV-2
Whole genome sequencing-based comparisons of the 2003
Severe Acute Respiratory Syndrome Coronavirus (SARS-CoV) and the 2019
SARS-CoV-2 revealed conserved receptor binding domain (RBD) and host
cell receptor, angiotensin-converting enzyme 2 (ACE2). In line with
this, the authors tested cross-reactivity of murine monoclonal
antibodies (mAbs) previously generated against the SARS-CoV spike (S)
glycoprotein involved in viral entry. One of the screened mAb, 1A9, was
able to bind and cross-neutralize multiple strains of SARS-CoV, as well
as, detect the S protein in SARS-CoV-2-infected cells. mAb 1A9 was
generated using an immunogenic fragment in the S2 subunit of SARS-CoV
and binds through a novel epitope within the S2 subunit at amino acids
1111-1130. It is important to note that CD8+ T lymphocyte epitopes
overlap with these residues, suggesting that S2 subunit could be
involved in inducing both, humoral and cell-mediated immunity.
13.49.3 Limitations
The authors used previously generated mouse mAbs
against the S protein in SARS-CoV expressed in mammalian cell line.
Future experimental validation using COVID-19 patient samples is needed
to validate these findings. In addition, the results of these studies
are predominantly based on in vitro experiments and so, evaluating the
effects of the mAb 1A9 in an animal model infected with this virus will
help us better understand the host immune responses in COVID-19 and
potential therapeutic vaccines.
13.49.4 Significance
This study identified mAbs that recognize the new
coronavirus, SARS-Cov-2. These cross-reactive mAbs will help in
developing diagnostic assays for COVID-19.
13.49.5 Credit
This review was undertaken by Tamar Plitt and Katherine Lindblad as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.50 Mortality of COVID-19 is Associated with Cellular Immune Function Compared to Immune Function in Chinese Han Population
Retrospective study of the clinical characteristics of 752
patients infected with COVID-19 at Chinese PLA General Hospital, Peking
Union Medical College Hospital, and affiliated hospitals at Shanghai
University of medicine & Health Sciences. This study is the first one
that compares PB from healthy controls from the same regions in Shanghai
and Beijing, and infected COVID-19 patients to standardize a reference
range of WBCs of people at high risk.
13.50.3 Limitations
Lower levels of leukocyte counts -B cells, CD4 and CD8 T
cells- correlated with mortality (WBCs are significantly lower in severe
or critical UCI patients vs mild ones). Based on 14,117 normal controls
in Chinese Han population (ranging in age from 18-86) it is recommended
that reference ranges of people at high risk of COVID-19 infection are
CD3+ lymphocytes below 900 cells/mm3, CD4+ lymphocytes below 500
cells/mm3, and CD8+ lymphocytes below 300 cells/mm3. Importantly, this
study also reported that the levels of D-dimer, C-reactive protein and
IL-6 were elevated in COVID-19 pts., indicating clot formation, severe
inflammation and cytokine storm.
13.50.4 Significance
This study sets a threshold to identify patients at risk
by analyzing their levels of leukocytes, which is an easy and fast
approach to stratify individuals that require hospitalization. Although
the study is limited (only counts of WBC are analyzed and not its
profile) the data is solid and statistically robust to correlate levels
of lymphopenia with mortality.
13.50.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.51 Retrospective Analysis of Clinical Features in 101 Death Cases with COVID-19
This is a retrospective study involving 101 death cases with COVID-19 in
Wuhan Jinyintan Hospital. The aim was to describe clinical,
epidemiological and laboratory features of fatal cases in order to
identify the possible primary mortality causes related to COVID-19.
Among 101 death cases, 56.44% were confirmed by RT-PCR and 43.6% by
clinical diagnostics. Males dominated the number of deaths and the
average age was 65.46 years. All patients died of respiratory failure
and multiple organs failure, except one (acute coronary syndrome). The
predominant comorbidities were hypertension (42.57%) and diabetes
(22.77%). 25.74% of the patients presented more than two underlying
diseases. 82% of patients presented myocardial enzymes abnormalities at
admission and further increase in myocardial damage indicators with
disease progression: patients with elevated Troponin I progressed faster
to death. Alterations in coagulation were also detected. Indicators of
liver and kidney damage increased 48 hours before death. The authors
studied the deceased patients’ blood type and presented the following
results: type A (44.44%), type B (29.29%), type AB (8.08%) and type O
(18.19%), which is inconsistent with the distribution in Han population
in Wuhan.
Clinical analysis showed that the most common symptom was fever (91.9%),
followed by cough and dyspnea. The medium time from onset of symptoms to
acute respiratory distress syndrome (ARDS) development was 12 days.
Unlike SARS, only 2 patients with COVID-19 had diarrhea. 98% presented
abnormal lung imaging at admission and most had double-lung
abnormalities. Related to the laboratorial findings some inflammatory
indicators gradually increased during the disease progression, such as
IL-6 secretion in the circulation, procalcitonin (PCT) and C-reactive
protein (CRP), while platelets numbers decreased. The authors also
reported an initial lymphopenia that was followed by an increase in the
lymphocytes numbers. Neutrophil count increased with disease
progression.
The patients received different treatments such as antiviral drugs
(60.40%), glucocorticoids, thymosin and immunoglobulins. All patients
received antibiotic treatment and some received antifungal drugs. All
patients received oxygen therapy (invasive or non-invasive ones).
13.51.3 Limitations
This study involves just fatal patients, lacking comparisons with other
groups of patients e.g. patients that recovered from COVID-19. The
authors didn’t discuss the different approaches used for treatments and
how these may affect the several parameters measured. The possible
relationship between the increase of inflammatory indicators and
morbidities of COVID-19 are not discussed.
13.51.4 Significance
This study has the largest cohort of fatal cases reported so far. The
authors show that COVID-19 causes fatal respiratory distress syndrome
and multiple organ failure. This study highlights prevalent myocardial
damage and indicates that cardiac function of COVID-19 patients should
be carefully monitored. The data suggest that Troponin I should be
further investigated as an early indicator of patients with high risk of
accelerated health deterioration. Secondary bacterial and fungal
infections were frequent in critically ill patients and these need to be
carefully monitored in severe COVID-19 patients. Differences in blood
type distribution were observed, suggesting that type A is detrimental
while type O is protective – but further studies are needed to confirm
these findings and elucidate if blood type influences infection or
disease severity. Several inflammatory indicators (neutrophils, PCT, CRP
and IL-6, D-dimer) increased according to disease severity and should be
assessed as biomarkers and to better understand the biology of
progression to severe disease.
13.51.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.52 Relationship between the ABO Blood Group and the COVID-19 Susceptibility
These authors compared the ABO blood group of 2,173 patients with
RT-PCR-confirmed COVID-19 from hospitals in Wuhan and Shenzhen with the
ABO blood group distribution in unaffected people in the same cities
from previous studies (2015 and 2010 for Wuhan and Shenzhen,
respectively). They found that people with blood group A are
statistically over-represented in the number of those infected and who
succumb to death while those with blood group O are statistically
underrepresented with no influence of age or sex.
13.52.3 Limitations
This study compares patients with COVID-19 to the general population
but relies on data published 5 and 10 years ago for the control. The
mechanisms that the authors propose may underlie the differences they
observed require further study.
13.52.4 Significance
Risk stratification based on blood group may be beneficial for
patients and also healthcare workers in infection control. Additionally,
investigating the mechanism behind these findings could lead to better
developing prophylactic and therapeutic targets for COVID-19.
13.52.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.53 The inhaled corticosteroid ciclesonide blocks coronavirus RNA replication by targeting viral NSP15
This study reconsiders the use of inhaled corticosteroids in the
treatment of pneumonia by coronavirus. Corticosteroids were associated
with increased mortality for SARS in 2003 and for MERS in 2013, probably
due to that fact that systemic corticosteroids suppress the innate
immune system, resulting in increased viral replication. However, some
steroid compounds might block coronavirus replication. The authors
screened steroids from a chemical library and assessed the viral growth
suppression and drug cytotoxicity. Ciclesonide demonstrated low
cytotoxicity and potent suppression of MERS-CoV viral growth. The
commonly used systemic steroids cortisone, prednisolone and
dexamethasone did not suppress viral growth, nor did the commonly used
inhaled steroid fluticasone. To identify the drug target of virus
replication, the authors conducted 11 consecutive MERS-CoV passages in
the presence of ciclesonide or mometasone, and they could generate a
mutant virus that developed resistance to ciclesonide, but not to
mometasone. Afterwards, they performed next-generation sequencing and
identified an amino acid substitution in nonstructural protein 15
(NSP15) as the predicted mechanism for viral resistance to ciclesonide.
The authors were able to successfully generate a recombinant virus
carrying that amino acid substitution, which overcome the antiviral
effect of ciclesonide, suggesting that ciclosenide interacts with NSP15.
The mutant virus was inhibited by mometasone, suggesting that the
antiviral target of mometasone is different from that of ciclesonide.
Lastly, the effects of ciclesonide and mometason on suppressing the
replication of SARS-CoV-2 were evaluated. Both compounds were found to
suppress viral replication with a similar efficacy to lopinavir.
13.53.3 Limitations
Most of the experiments, including the identification of the mutation in
NSP15 were conducted with MERS-CoV. This is not the closest related
virus to SARS-CoV-2, as that would be SARS-CoV. Thus, to repeat the
initial experiments with SARS-CoV, or preferably SARS-CoV-2, is
essential. The manuscript should address this and, therefore, it will
require considerable editing for organization and clarity. Also, in
terms of cell immunogenic epitopes, while SARS-CoV-2 spike protein
contains several predicted B and T cell immunogenic epitopes that are
shared with other coronaviruses, some studies have shown critical
differences between MERS-CoV, SARS-CoV and SARS-CoV-2. A main criticism
is that the authors only used VeroE6/TMPRSS2 cells to gauge the direct
cytotoxic effects of viral replication. To evaluate this in other cell
lines, including human airway epithelial cells, is crucial, as the
infectivity of coronavirus strains greatly varies in different cell
lines,
13.53.4 Significance
Nevertheless, these findings encourage evaluating ciclesonide and
mometasone as better options for patients with COVID-19 in need of
inhaled steroids, especially as an alternative to other corticosteroids
that have been shown to increase viral replication in vitro. This should
be evaluated in future clinical studies.
13.53.5 Credit
This review was undertaken by Alvaro Moreira, MD as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.54 A human monoclonal antibody blocking SARS-CoV-2 infection **
The authors reported a human monoclonal
antibody that neutralizes SARS-CoV-2 and SARS-Cov which belong to same
family of corona viruses. For identifying mAbs, supernatants of a
collection of 51 hybridomas raised against the spike protein of SARS-CoV
(SARS-S) were screened by ELISA for cross-reactivity against the spike
protein of SARS-CoN2 (SARS2-S). Hybridomas were derived from immunized
transgenic H2L2 mice (chimeric for fully human VH-VL and rat constant
region). Four SARS-S hybridomas displayed cross-reactivity with SARS2-S,
one of which (47D11) exhibited cross-neutralizing activity for SARS-S
and SARS2-S pseudotyped VSV infection. A recombinant, fully human IgG1
isotype antibody was generated and used for further characterization.
The humanized 47D11 antibody inhibited infection of VeroE6 cells with
SARS-CoV and SARS-CoV-2 with IC50 values of 0.19 and 0.57 μg/ml
respectively. 47D11 mAb bound a conserved epitope on the spike receptor
binding domain (RBD) explaining its ability to cross-neutralize SARS-CoV
and SARS-CoV-2. 47D11 was shown to target the S1B RBD of SARS-S and
SARS2-S with similar affinities. Interestingly, binding of 47D11 to
SARS-S1B and SARS2-S1B did not interfere with S1B binding to ACE2
receptor-expressing cells assayed by flow cytometry.
13.54.3 Limitations
These results show that the human 47D11 antibody
neutralizes SARS-CoV and SARS-Cov2 infectivity via an as yet unknown
mechanism that is different from receptor binding interference.
Alternative mechanisms were proposed but these as yet remain to be
tested in the context of SARS-CoV2. From a therapeutic standpoint and in
the absence of in vivo data, it is unclear whether the 47D11 ab can
alter the course of infection in an infected host through virus
clearance or protect an uninfected host that is exposed to the virus.
There is a precedent for the latter possibility as it relates to
SARS-CoV that was cited by the authors and could turn out to be true for
SARS-CoV2.
13.54.4 Significance
This study enabled the identification of novel neutralizing
antibody against COV-that could potentially be used as first line of
treatment in the near future to reduce the viral load and adverse
effects in infected patients. In addition, neutralizing antibodies such
as 47D11 represent promising reagents for developing
antigen-antibody-based detection test kits and assays.
13.54.5 Credit
This review was edited by K. Alexandropoulos as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
Heat inactivation of serum interferes with the immunoanalysis of
antibodies to SARS-CoV-2
The use of heat inactivation to neutralize pathogens in serum samples
collected from suspected COVID-19 patients reduces the sensitivity of a
fluorescent immunochromatographic assay to detect anti-SARS-CoV-2 IgM
and IgG.
Major findings
Coronaviruses can be killed by heat inactivation, and this is an
important safety precaution in laboratory manipulation of clinical
samples. However, the effect of this step on downstream
SARS-CoV-2-specific serum antibody assays has not been examined. The
authors tested the effect of heat inactivation (56 deg C for 30 minutes)
versus no heat inactivation on a fluorescence immunochromatography
assay. Heat inactivation reduced all IgM measurements by an average of
54% and most IgG measurements (22/36 samples, average reduction of 50%),
consistent with the lower thermal stability of IgM than that of IgG.
Heat inactivation caused a subset of IgM but not IgG readings to fall
below a specified positivity threshold.
Limitations
Limitations included the use of only one type of assay for testing heat
inactivated vs non-inactivated sera, and the use of the same baseline
for heat inactivated and non-inactivated sera. The results indicate that
heat inactivation affects the quantification of SARS-CoV-2-antibody
response, specially IgM, but still allows to distinguish positive
specific IgG. Therefore, the effect of heat inactivation should be
studied when designing assays that quantitatively associate
immunoglobulin levels (especially IgM) to immune state.
Review by Andrew M. Leader as part of a project by students, postdocs
and faculty at the Immunology Institute of the Icahn school of medicine,
Mount Sinai.
13.55 Immune phenotyping based on neutrophil-to-lymphocyte ratio and IgG predicts disease severity and outcome for patients with COVID-19
In a cohort of 222 patients, anti-SARS-CoV-2 IgM and IgG levels were
analyzed during acute and convalescent phases (up to day 35) and
correlated to the diseases’ severity. The same was done with
neutrophil-to-lymphocyte ratio. High IgG levels and high
neutrophil-to-lymphocyte ratio in convalescence were both independently
associated to the severity of the disease. The simultaneous occurrence
of both of these laboratory findings correlated even stronger to the
diseases’ severity.
Severe cases with high neutrophil-to-lymphocyte ratios had clearly
higher levels of IL-6. The authors propose that a robust IgG response
leads to immune-mediated tissue damage, thus explaining the worse
outcome in patients with overexuberant antibody response.
13.55.3 Limitations
A main criticism is that the criteria for stratifying patients in severe
vs. non-severe are not described. The only reference related to this is
the difference between the percentage of patients who needed mechanical
ventilation, which was greater in patients with both high IgG levels and
high neutrophil-to-lymphocyte ratio. No patient with both low IgG levels
and low neutrophil-to-lymphocyte ratio was treated with mechanical
ventilation.
The proposed correlation of severity with IL-2 and IL-10 levels is not
very strong.
Furthermore, although mostly ignored in the paper’s discussion, one of
the most interesting findings is that an early increase in
anti-SARS-CoV-2 IgM levels also seems to correlate with severe disease.
However, as only median values are shown for antibody kinetics curves,
the extent of variation in acute phase cannot be assessed.
13.55.4 Significance
Anti-SARS-CoV-2 IgG levels and with neutrophil-to-lymphocyte ratio
predict severity of COVID-19 independently of each other. An additive
predictive value of both variables is noticeable. Importantly, an
early-on increase in anti-SARS-CoV-2 IgM levels also seem to predict
outcome.
13.55.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.56 Reinfection could not occur in SARS-2 CoV-2 infected rhesus macaques
This study addresses the issue or acquired immunity after a primary
COVID-19 infection in rhesus monkeys. Four Chinese rhesus macaques were
intratracheally infected with SARS-CoV-2 and two out of the four were
re-infected at 28 days post initial infection (dpi) with the same viral
dose after confirming the recovery by the absence of clinical symptoms,
radiological abnormalities and viral detection (2 negative RT-PCR
tests). While the initial infection led the viral loads in nasal and
pharyngeal swabs that reach approximately 6.5 log10 RNA copies/ml at 3
dpi in all four monkeys, viral loads in the swabs tested negative after
reinfection in the two reinfected monkeys. In addition, the necropsies
from a monkey (M1) at 7 days after primary infection, and another monkey
(M3) at 5 days post reinfection, revealed the histopathological damages
and viral replication in the examined tissues from M1, while no viral
replication as well as no histological damages were detected in the
tissues from M3. Furthermore, sera from three monkeys at 21 and 28 dpi
exhibited neutralizing activity against SARS-CoV-2 in vitro, suggesting
the production of protective neutralizing antibodies in these monkeys.
Overall, this study indicates that primary infection with SARS-CoV-2 may
protect from subsequent exposure to the same virus.
13.56.3 Limitations
In human, virus has been detected by nasopharyngeal swabs until 9 to 15
days after the onset of symptoms. In the infected monkeys in this study,
virus were detected from day 1 after the infection, declining to
undetectable level by day 15 post infection. It may suggest that there
is a faster viral clearance mechanism in monkeys, therefore the
conclusions of reinfection protection for humans need to be carefully
considered. In addition, only two monkeys were re-infected in this study
and the clinical signs of these monkeys were not similar: M3 did not
show weight loss and M4 showed relatively higher fever on the day of
infection and the day of re-challenge.
13.56.4 Significance
This study showed clear viral clearance and no indications of relapse or
viremia after a secondary infection with SARS-CoV-2 in a Chinese rhesus
macaque model. These results support the idea that patients with full
recovery (two negative RT-PCR results) may also be protected from
secondary SARS-CoV-2 infection. Recovered patients may be able to
reintegrate to normal public life and provide protective serum perhaps
even if having had a mild infection. The results are also encouraging
for successful vaccine development against SARS-CoV-2.
13.56.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.57 A highly conserved cryptic epitope in the receptor-binding domains of SARS-CoV-2 and SARS-CoV
Given the sequence similarity of the surface spike glycoprotein (S) of
SARS-CoV-2 and SAR-CoV, Yuan et al. (2020) propose that neutralizing
antibodies isolated from convalescent SARS-CoV patients may offer
insight into cross-reactive antibodies targeting SARS-CoV-2. In
particular, they find that the receptor-binding domain (RBD) of
SARS-CoV-2 S protein shares 86% sequence similarity with the RBD of
SARS-CoV S protein that binds to the CR3022 neutralizing antibody.
CR3022 also displays increased affinity for the “up” conformation of the
SARS-CoV-2 S protein compared to the “down” conformation as it does for
the SARS-CoV S protein. Therefore, the authors propose that this
cross-reactive antibody may confer some degree of protection in vivo
even if it fails to neutralize in vitro.
13.57.3 Limitations
Although the authors offer a logical rationale for
identifying cross-reactive neutralizing antibodies derived from
SARS-CoV, their study using only CR3022 failed to demonstrate whether
this approach will be successful. After all, CR3022 failed to neutralize
in vitro despite the binding affinity to a similar epitope on
SARS-CoV-2. They would benefit from testing more candidates and using an
in vivo model to demonstrate their claim that protection may be
possible in the absence neutralization if combinations are used in
vivo.
13.57.4 Significance
The ability to make use of previously characterized
neutralizing antibodies for conserved epitopes can expedite drug design
and treatment options.
13.57.5 Credit
This review was undertaken by Dan Fu Ruan, Evan Cody and Venu Pothula as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.58 Highly accurate and sensitive diagnostic detection of SARS-CoV-2 by digital PCR
The authors present a digital PCR (dPCR) diagnostic test for
SARS-CoV-2 infection. In 103 individuals that were confirmed in a
follow-up to be infected, the standard qPCR test had a positivity
rate of 28.2% while the dPCR test detected 87.4% of the infections
by detecting an additional 61 positive cases. The authors also
tested samples from close contacts (early in infection stage) and
convalescing individuals (late in infection stage) and were able to
detect SARS-CoV-2 nucleic acid in many more samples using dPCR
compared to qPCR.
13.58.3 Limitations
I did not detect limitations.
13.58.4 Significance
The authors make a strong case for the need for a highly sensitive
and accurate confirmatory method for diagnosing COVID-19 during this
outbreak and present a potential addition to the diagnostic arsenal.
They propose a dPCR test that they present has a dramatically lower
false negative rate than the standard RT-qPCR tests and can be
especially beneficial in people with low viral load, whether they
are in the earlier or later stages of infection.
13.58.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.59 SARS-CoV-2 invades host cells via a novel route: CD147-spike protein
The authors propose a novel mechanism of SARS-CoV-2 viral entry through
the interaction of the viral spike protein (SP) and the immunoglobulin
superfamily protein CD147 (also known as Basigin). Using an in-house
developed humanized antibody against CD147 (maplazumab), they show that
blocking CD147 decreases viral replication in Vero E6 cells. Using
surface plasmon resonance (SPR), ELISA, and Co-IP assays, they show that
the spike protein of SARS-CoV-2 directly interacts with CD147. Lastly,
they utilize immune-election microscopy to show spike protein and CD147
localize to viral inclusion bodies of Vero E6 cells.
13.59.3 Limitations
The authors claim that an anti-CD147 antibody (Meplazumab) inhibits
SARS-CoV-2 replication by testing cell growth and viral load in cells
infected with SARS-CoV-2, however there are key pieces of this
experiment that are missing. First, the authors fail to use a
non-specific antibody control. Second, the authors claim that viral
replication is inhibited, and that they test this by qPCR, however this
data is not shown. To further prove specificity, the authors should
introduce CD147 to non-susceptible cells and show that they become
permissive.
The authors claim that there is a direct interaction between CD147 and
SP through SPR, ELISA, and Co-IP, and this data seems generally
convincing. The electron microscopy provides further correlative
evidence that SARS-CoV-2 may interact with CD147 as they are both found
in the same viral inclusion body. A quantification of this data would
make the findings more robust.
Finally, the data in this paper lacks replicates, error bars, and
statistics to show that the data are reproducible and statistically
significant.
13.59.4 Significance
It has been shown in various studies that SARS-CoV-2 binds to the cell
surface protein ACE2 for cell entry, yet ACE2 is highly expressed in
heart, kidney, and intestinal cells, raising the concern that blocking
ACE2 would result in harmful side effects [1319] CD147 on the other hand
is highly expressed in various tumor types, inflamed tissues, and
pathogen infected cells, suggesting that the inhibition of CD147 would
not result in major side effects [1320,1321] The research in this paper has
resulted in an ongoing clinical trail in China to test the safety and
efficacy of anti-CD147 Meplazumab to treat COVID-19. (ClinicalTrails.gov
identifier NCT04275245).
13.59.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.60 Blood single cell immune profiling reveals that interferon-MAPK pathway mediated adaptive immune response for COVID-19
The authors performed single-cell RNA sequencing (scRNAseq) of
peripheral blood mononuclear cells isolated from whole blood samples of
COVID-19 patients (n=10). Data was compared to scRNAseq of samples
collected from patients with influenza A (n=1), acute pharyngitis (n=1),
and cerebral infarction (n=1), as well as, three healthy controls.
COVID-19 patients were categorized into those with moderate (n=6),
severe (n=1), critical (n=1), and cured (n=2) disease. Analysis across
all COVID-19 disease levels revealed 56 different cellular subtypes,
among 17 immune cell types; comparisons between each category to the
normal controls revealed increased proportions of CD1c+ dendritic
cells, CD8+ CTLs, and plasmacytoid dendritic cells and a decrease in
proportions of B cells and CD4+ T cells.
TCR sequencing revealed that greater clonality is associated with
milder COVID-19 disease; BCR sequencing revealed that COVID-19
patients have circulating antibodies against known viral antigens,
including EBV, HIV, influenza A, and other RNA viruses. This may suggest
that the immune response to SARS-CoV-2 infection elicits production of
antibodies against known RNA viruses.
Excluding enriched pathways shared by COVID-19 patients and patients
with other conditions (influenza A, acute pharyngitis, and cerebral
infarction), the authors identified the interferon-MAPK signaling
pathway as a major response to SARS-CoV-2 infection. The authors
performed quantitative real-time reverse transcriptase polymerase chain
reaction (RT-PCR) for interferon-MAPK signaling genes: IRF27, BST2,
and FOS. These samples were collected from a separate cohort of
COVID-19 patients (critical, n=3; severe, n=3; moderate, n=19; mild,
n=3; and cured, n=10; and healthy controls, n=5). Notably, consistent
with the original scRNAseq data, FOS showed up-regulation in COVID-19
patients and down-regulation in cured patients. The authors propose
that FOS may be a candidate marker gene for curative COVID-19
disease.
13.60.3 Limitations
The sample size of this study is limited. To further delineate
differences in the immune profile of peripheral blood of COVID-19
patients, a greater sample size is needed, and longitudinal samples are
needed, as well. A better understanding of the immunological
interactions in cured patients, for example, would require a profile
before and after improvement.
Moreover, the conclusions drawn from this scRNAseq study point to
potential autoimmunity and immune deficiency to distinguish different
severities of COVID-19 disease. However, this requires an expanded
number of samples and a more robust organization of specific immune cell
subtypes that can be compared across different patients. Importantly,
this criterion is likely needed to ensure greater specificity in
identifying markers for COVID-19 infection and subsequent immune
response.
13.60.4 Significance
At the single-cell level, COVID-19 disease has been characterized in the
lung, but a greater understanding of systemic immunological responses is
furthered in this study. Type I interferon is an important signaling
molecule for the anti-viral response. The identification of the
interferon-MAPK signaling pathway and the differential expression of
MAPK regulators between patients of differing COVID-19 severity and
compared to cured patients may underscore the importance of either
immune deficiency or autoimmunity in COVID-19 disease.
13.60.5 Credit
This review was undertaken by Matthew D. Park as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.61 Cross-reactive antibody response between SARS-CoV-2 and SARS-CoV infection.
The authors explore the antigenic differences between SARS-CoV-2 and
SARS-CoV by analyzing plasma samples from SARS-CoV-2 (n = 15) and
SARS-CoV (n = 7) patients. Cross-reactivity in antibody binding to the
spike protein between SARS-CoV-2 and SARS-CoV was found to be common,
mostly targeting non-RBD regions in plasma from SARS-CoV-2 patients.
Only one SARS-CoV-2 plasma sample was able to cross-neutralize SARS-CoV,
with low neutralization activity. No cross-neutralization response was
detected in plasma from SARS-CoV patients.
To further investigate the cross-reactivity of antibody responses to
SARS-CoV-2 and SARS-CoV, the authors analyzed the antibody response of
plasma collected from mice infected or immunized with SARS-CoV-2 or
SARS-CoV (n = 5 or 6 per group). Plasma from mice immunized with
SARS-CoV-2 displayed cross-reactive responses to SARS-CoV S ectodomain
and, to a lesser extent, SARS-CoV RBD. Similarly, plasma from mice
immunized with SARS-CoV displayed cross-reactive responses to SARS-CoV-2
S ectodomain. Cross-neutralization activity was not detected in any of
the mouse plasma samples.
13.61.3 Limitations
The size of each patient cohort is insufficient to accurately determine
the frequency of cross-reactivity and cross-neutralization in the
current SARS-CoV-2 pandemic. Recruitment of additional patients from a
larger range of geographical regions and time points would also enable
exploration into the effect of the genetic diversity and evolution of
the SARS-CoV-2 virus on cross-reactivity. This work would also benefit
from the mapping of specific epitopes for each sample. Future studies
may determine whether the non-neutralizing antibody responses can confer
in vitro protection or lead to antibody-dependent disease enhancement.
13.61.4 Significance
The cross-reactive antibody responses to S protein in the majority of
SARS-CoV-2 patients is an important consideration for development of
serological assays and vaccine development during the current outbreak.
The limited extent of cross-neutralization demonstrated in this study
indicates that vaccinating to cross-reactive conserved epitopes may have
limited efficacy, presenting a key concern for the development of a more
universal coronavirus vaccine to address the global health risk of novel
coronavirus outbreaks.
13.61.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.62 The feasibility of convalescent plasma therapy in severe COVID-19 patients: a pilot study
This is the first report to date of convalescent plasma therapy as a
therapeutic against COVID-19 disease. This is a feasibility pilot study.
The authors report the administration and clinical benefit of 200 mL of
convalescent plasma (CP) (1:640 titer) derived from recently cured
donors (CP selected among 40 donors based on high neutralizing titer and
ABO compatibility) to 10 severe COVID-19 patients with confirmed
viremia. The primary endpoint was the safety of CP transfusion. The
secondary endpoint were clinical signs of improvement based on symptoms
and laboratory parameters.
The authors reported use of methylene blue photochemistry to inactivate
any potential residual virus in the plasma samples, without compromising
neutralizing antibodies, and no virus was detected before transfusion.
The authors report the following:
No adverse events were observed in all patients, except 1 patient
who exhibited transient facial red spotting.
All patients showed significant improvement in or complete
disappearance of clinical symptoms, including fever, cough,
shortness of breath, and chest pain after 3 days of CP therapy.
Reduction of pulmonary lesions revealed by chest CT.
Elevation of lymphocyte counts in patients with lymphocytopenia.
Increase in SaO2 in all patients, indicative of recuperating lung
function.
Resolution of SARS-CoV-2 viremia in 7 patients and increase in
neutralizing antibody titers in 5 patients. Persistence of
neutralizing antibody levels in 4 patients.
13.62.3 Limitations
It is important to note that most recipients had high neutralization
titers of antibodies before plasma transfusion and even without
transfusion it would be expected to see an increase in neutralizing
antibodies over time. In addition to the small sample set number (n=10),
there are additional limitations to this pilot study:
All patients received concurrent therapy, in addition to the CP
transfusion. Therefore, it is unclear whether a combinatorial or
synergistic effect between these standards of care and CP
transfusion contributed to the clearance of viremia and improvement
of symptoms in these COVID-19 patients.
The kinetics of viral clearance was not investigated, with respect
to the administration of CP transfusion. So, the definitive impact
of CP transfusion on immune dynamics and subsequent viral load is
not well defined.
Comparison with a small historical control group is not ideal.
13.62.4 Significance
For the first time, a pilot study provides promising results involving
the use of convalescent plasma from cured COVID-19 patients to treat
others with more severe disease. The authors report that the
administration of a single, high-dose of neutralizing antibodies is
safe. In addition, there were encouraging results with regards to the
reduction of viral load and improvement of clinical outcomes. It is,
therefore, necessary to expand this type of study with more
participants, in order to determine optimal dose and treatment kinetics.
It is important to note that CP has been studied to treat H1N1
influenza, SARS-CoV-1, and MERS-CoV, although it has not been proven to
be effective in treating these infections.
13.62.5 Credit
Review by Matthew D. Park and revised by Alice O. Kamphorst and Maria A. Curotto de Lafaille as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.63 Hydroxychloroquine and azithromycin as a treatment of COVID-19: results of an open label non-randomized clinical trial
This study was a single-arm, open label clinical trial with
600 mg hydroxychloroquine (HCQ) in the treatment arm (n = 20). Patients
who refused participation or patients from another center not treated
with HCQ were included as negative controls (n = 16). Among the patients
in the treatment arm, 6 received concomitant azithromycin to prevent
superimposed bacterial infection. The primary endpoint was respiratory
viral loads on day 6 post enrollment, measured by nasopharyngeal swab
followed by real-time reverse transcription-PCR.
HCQ alone was able to significantly reduce viral loads by day 6 (n =
8/14, 57.1% complete clearance, p < 0.001); azithromycin appears to
be synergistic with HCQ, as 6/6 patients receiving combined treatment
had complete viral clearance (p < 0.001).
13.63.3 Limitations
Despite what is outlined above, this study has a number
of limitations that must be considered. First, there were originally n =
26 patients in the treatment arm, with 6 lost to follow up for the
following reasons: 3 transferred to ICU, 1 discharge, 1
self-discontinued treatment d/t side effects, and 1 patient expired.
Total length of clinical follow up was 14 days, but the data beyond day
6 post-inclusion are not shown.
Strikingly, in supplementary table 1, results of the real-time RT-PCR
are listed for the control and treatment arms from D0 – D6. However, the
data are not reported in a standard way, with a mix of broadly positive
or negative result delineation with Ct (cycle threshold) values, the
standard output of real time PCR. It is impossible to compare what is
defined as a positive value between the patients in the control and
treatment arms without a standardized threshold for a positive test.
Further, the starting viral loads reported at D0 in the groups receiving
HCQ or HCQ + azithromycin were significantly different (ct of 25.3 vs
26.8 respectively), which could explain in part the differences observed
in the response to treatment between 2 groups. Finally, patients in the
control arm from outside the primary medical center in this study
(Marseille) did not actually have samples tested by PCR daily. Instead,
positive test results from every other day were extrapolated to mean
positive results on the day before and after testing as well (Table 2,
footnote a).
Taken together, the results of this study suggest that HCQ represents a
promising treatment avenue for COVID-19 patients. However, the limited
size of the trial, and the way in which the results were reported does
not allow for other medical centers to extrapolate a positive or
negative result in the treatment of their own patients with HCQ +/-
azithromycin. Further larger randomized clinical trials will be required
to ascertain the efficacy of HCQ +/- azithromycin in the treatment of
COVID-19.
13.63.4 Significance
Chloroquine is thought to inhibit viral infection,
including SARS-Cov-2, by increasing pH within endosomes and lysosomes,
altering the biochemical conditions required for viral fusion [439,1326].
However, chloroquine also has immuno-modulatory effects that I think may
play a role. Chloroquine has been shown to increase CTLA-4 expression at
the cell surface by decreasing its degradation in the endo-lysosome
pathway; AP-1 traffics the cytoplasmic tail of CTLA-4 to lysosomes, but
in conditions of increased pH, the protein machinery required for
degradation is less functional [1327]. As such, more CTLA-4 remains in
endosomes and is trafficked back to the cell surface. It is possible
that this may also contribute to patient recovery via reduction of
cytokine storm, in addition to the direct anti-viral effects of HCQ.
13.63.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.64 Recapitulation of SARS-CoV-2 Infection and Cholangiocyte Damage with Human Liver Organoids
Used human liver ductal organoids to determine ACE2+ cholangiocytes
in healthy liver (2.92% of all cells) are infectable and support
SARS-CoV-2 viral replication.
Plaque-purified SARS-CoV-2 viral infection disrupted organoid
barrier and bile transporting functions of cholangiocytes through
dysregulation of genes involved in tight junction formation (CLDN1)
and bile acid transportation (ASBT and CFTR).
13.64.3 Limitations:
Unclear if liver damage observed in patients due to direct
cholangiocyte infection or due to secondary immune/cytokine effects.
This study argues for direct damage as it lacks immune contexture;
but further studies needed with autopsy samples or organoid-immune
cell co-culture to conclude strongly.
Would be important to measure cholangiocyte-intrinsic anti-viral
response and alarmins secreted upon infection, and furthermore study
tropism of various immune cells to conditioned media from organoids
infected with SARS-CoV-2.
Does not address how cirrhotic liver or
alcohol/smoking/obesity-associated liver organoids respond to
SARS-CoV-2 infectivity and replication, worth pursuing to
experimentally address clinical data indicating co-morbidities.
13.64.4 Significance
Useful model to rapidly study drug activity against SARS-CoV-2
infection in liver, while monitoring baseline liver damage.
Liver abnormality observed in >50% of CoVID-19 patients; the
results from this study could explain the bile acid accumulation and
consequent liver damage observed.
13.64.5 Credit
Review by Samarth Hegde as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.65 The sequence of human ACE2 is suboptimal for binding the S spike protein of SARS coronavirus 2
Severe Acute Respiratory Syndrome Coronavirus 2
(SARS-CoV-2) infects cells through S spike glycoprotein binding
angiotensin-converting enzyme (ACE2) on host cells. S protein can bind
both membrane-bound ACE2 and soluble ACE2 (sACE2), which can serve as a
decoy that neutralizes infection. Recombinant sACE2 is now being tested
in clinical trials for COVID-19. To determine if a therapeutic sACE2
with higher affinity for S protein could be designed, authors generated
a library containing every amino acid substitution possible at the 117
sites spanning the binding interface with S protein. The ACE2 library
was expressed in human Expi293F cells and cells were incubated with
medium containing the receptor binding domain (RBD) of SARS-CoV-2 fused
to GFP. Cells with high or low affinity mutant ACE2 receptor compared to
affinity of wild type ACE2 for the RBD were FACS sorted and transcripts
from these sorted populations were deep sequenced. Deep mutagenesis
identified numerous mutations in ACE2 that enhance RBD binding. This
work serves to identify putative high affinity ACE2 therapeutics for the
treatment of CoV-2.
13.65.3 Limitations
The authors generated a large library of mutated
ACE2, expressed them in human Expi293F cells, and performed deep
mutagenesis to identify enhanced binders for the RBD of SARS-CoV-2 S
protein. While these data serve as a useful resource, the ability of the
high affinity ACE2 mutants identified to serve as therapeutics needs
further validation in terms of conformational stability when purified as
well as efficacy/safety both in vitro and in vivo. Additionally,
authors mentioned fusing the therapeutic ACE2 to Fc receptors to elicit
beneficial host immune responses, which would need further design and
validation.
13.65.4 Significance
This study identified structural ACE2 mutants that
have potential to serve as therapeutics in the treatment of SARS-CoV-2
upon further testing and validation.
13.65.5 Credit
This review was undertaken by Katherine Lindblad and Tamar Plitt as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
Title: A serological assay to detect SARS-Cov-2 seroconversion in
humans
Note: the authors of this review work in the same institution as the
authors of the study
Main findings:
Production of recombinant whole Spike (S) protein and the smaller
Receptor Binding Domain (RBD) based on the sequence of Wuhan-Hu-1
SARS-CoV-2 isolate. The S protein was modified to allow trimerization
and increase stability. The authors compared the antibody reactivity of
59 banked human serum samples (non-exposed) and 3 serum samples from
confirmed SARS-CoV-2 infected patients. All Covid-19 patient sera
reacted to the S protein and RBD domain compared to the control sera.
The authors also characterized the antibody isotypes from the Covid-19
patients, and observed stronger IgG3 response than IgG1. IgM and IgA
responses were also prevalent.
Limitations of the study:
The authors analyzed a total of 59 control human serum samples, and
samples from only three different patients to test for reactivity
against the RBD domain and full-length spike protein. It will be
important to follow up with a larger number of patient samples to
confirm the data obtained. Furthermore, it would be interesting to
assess people at different age groups and determine whether unexposed
control kids have a higher “background”.
Applications of the assay described in this study in diagnosis are
limited, since antibody response should start to be detectable only one
to two weeks after infection. Future studies will be required to assess
how long after infection this assay allow to detect anti-CoV2
antibodies. Finally, while likely, the association of seroconversion
with protective immunity against SARS-Cov-2 infection still needs to be
fully established.
Relevance:
This study has strong implications in the research against SARS-Cov-2.
First, it is now possible to perform serosurveys and determine who has
been infected, allowing a more accurate estimate of infection prevalence
and death rate. Second, if it is confirmed that re-infection does not
happen (or is rare), this assay can be used as a tool to screen
healthcare workers and prioritize immune ones to work with infected
patients. Third, potential convalescent plasma donors can now be
screened to help treating currently infected patients. Of note, this
assay does not involve live virus handling. experimentally, this is an
advantage as the assay does not require the precautions required by
manipulation of live virus. Finally, the recombinant proteins described
in this study represent new tools that can be used for further
applications, including vaccine development.
13.66 COMPARATIVE PATHOGENESIS OF COVID-19, MERS AND SARS IN A NON-HUMAN PRIMATE MODEL
This work assesses SARS-CoV-2 infection in young adult and aged
cynomolgus macaques. 4 macaques per age group were infected with
low-passage clinical sample of SARS-CoV-2 by intranasal and
intratracheal administration. Viral presence was assessed in nose,
throat and rectum through RT-PCR and viral culture. SARS-CoV-2
replication was confirmed in the respiratory track (including nasal
samples), and it was also detected in ileum. Viral nucleocapsid
detection by IHC showed infection of type I and II pneumocytes and
epithelia. Virus was found to peak between 2 and 4 days after
administration and reached higher levels in aged vs. young animals. The
early peak is consistent with data in patients and contrasts to SARS-CoV
replication. SARS-CoV-2 reached levels below detection between 8 and 21
days after inoculation and macaques established antibody immunity
against the virus by day 14. There were histopathological alteration in
lung, but no overt clinical signs. At day 4 post inoculation of
SARS-CoV-2, two of four animals presented foci of pulmonary
consolidation, with limited areas of alveolar edema and pneumonia, as
well as immune cell infiltration. In sum, cynomolgus macaques are
permissive to SARS- CoV-2 and develop lung pathology (less severe than
SARC-CoV, but more severe than MERS-CoV).
13.66.3 Limitations
Even though cynomolgus macaques were permissive to SARS-CoV-2
replication, it is unclear if the viral load reaches levels comparable
to humans and there wasn’t overt clinical pathology.
13.66.4 Significance
The development of platforms in which to carry out relevant
experimentation on SARS-CoV-2 pathophysiology is of great urgency.
Cynomolgus macaques offer an environment in which viral replication can
happen, albeit in a limited way and without translating into clinically
relevant symptoms. Other groups are contributing to SARS-CoV2 literature
using this animal model [1315], potentially showing protection against
reinfection in cured macaques. Therefore, this platform could be used to
examine SARS-CoV2 pathophysiology while studies in other animal models
are also underway [1254,1330].
13.66.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.67 Investigating the Impact of Asymptomatic Carriers on COVID-19 Transmission
Multiple studies reported the same level of infectiousness
between symptomatic and asymptomatic carriers of SARS-CoV-2. Given that
asymptomatic and undocumented carriers escape public health surveillance
systems, a better mathematical model of transmission is needed to
determine a more accurate estimate of the basic reproductive number
(R0) of the virus to assess the contagiousness of virus. The authors
developed a SEYAR dynamical model for transmission of the new
coronavirus that takes into account asymptomatic and undocumented
carriers. The model was validated using data reported from thirteen
countries during the first three weeks of community transmission. While
current studies estimate R0 to be around 3, this model indicates that
the value could range between 5.5 to 25.4.
13.67.3 Limitations
The SEYAR model realistically depicts
transmission of the virus only during the initial stages of the disease.
More data is necessary to better fit the model with current trends. In
addition, multiple factors (e.g. behavioral patterns, surveillance
capabilities, environmental and socioeconomic factors) affect
transmission of the virus and so, these factors must be taken into
consideration when estimating the R0.
13.67.4 Significance
Public health authorities use the basic reproductive
number to determine the severity of disease. An accurate estimate of
R0 will inform intervention strategies. This model can be applied to
different locations to assess the potential impact of COVID-19.
13.67.5 Credit
This review was undertaken by Tamar Plitt and Katherine Lindblad as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.68 Antibody responses to SARS-CoV-2 in COVID-19 patients: the perspective application of serological tests in clinical practice
This study investigated the profile of the acute antibody response
against SARS-CoV-2 and provided proposals for serologic tests in
clinical practice. Magnetic Chemiluminescence Enzyme Immunoassay was
used to evaluate IgM and IgG seroconversion in 285 hospital admitted
patients who tested positive for SARS-CoV-2 by RT-PCR and in 52 COVID-19
suspected patients that tested negative by RT-PCR. A follow up study
with 63 patients was performed to investigate longitudinal effects. In
addition, IgG and IgM titers were evaluated in a cohort of close
contacts (164 persons) of an infected couple.
The median day of seroconversion for both IgG and IgM was 13 days after
symptom onset. Patients varied in the order of IgM/ IgG seroconversion
and there was no apparent correlation of order with age, severity, or
hospitalization time. This led the authors to conclude that for
diagnosis IgM and IgG should be detected simultaneously at the early
phase of infection.
IgG titers, but not IgM titers were higher in severe patients compared
to non-severe patients after controlling for days post-symptom onset.
Importantly, 12% of COVID-19 patients (RT-PCR confirmed) did not meet
the WHO serological diagnosis criterion of either seroconversion or
> 4-fold increase in IgG titer in sequential samples. This suggests
the current serological criteria may be too stringent for COVID-19
diagnosis.
Of note, 4 patients from a group of 52 suspects (negative RT-PCR test)
had anti-SARS-Cov-2 IgM and IgG. Similarly, 4.3% (7/162) of “close
contacts” who had negative RT-PCR tests were positive for IgG and/or
IgM. This highlights the usefulness of a serological assay to identify
asymptomatic infections and/or infections that are missed by RT-PCR.
13.68.3 Limitations
This group’s report generally confirms the findings of others that have
evaluated the acute antibody response to SARS-Cov-2. However, these data
would benefit from inclusion of data on whether the participants had a
documented history of viral infection. Moreover, serum samples that were
collected prior to SARS-Cov-2 outbreak from patients with other viral
infections would serve as a useful negative control for their assay.
Methodological limitations include that only one serum sample per case
was tested as well as the heat inactivation of serum samples prior to
testing. It has previously been reported that heat inactivation
interferes with the level of antibodies to SARS-Cov-2 and their protocol
may have resulted in diminished quantification of IgM, specifically [1333].
13.68.4 Significance
Understanding the features of the antibody responses against SARS-CoV is
useful in the development of a serological test for the diagnosis of
COVID-19. This paper addresses the need for additional screening methods
that can detect the presence of infection despite lower viral titers.
Detecting the production of antibodies, especially IgM, which are
produced rapidly after infection can be combined with PCR to enhance
detection sensitivity and accuracy and map the full spread of infection
in communities, Moreover, serologic assays would be useful to screen
health care workers in order to identify those with immunity to care for
patients with COVID19.
13.68.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.69 SARS-CoV-2 specific antibody responses in COVID-19 patients
Antibodies specific to SARS-CoV-2 S protein, the S1 subunit and the RBD (receptor-binding domain) were detected in all SARS-CoV-2 patient sera by 13 to 21 days post onset of disease. Antibodies specific to SARS-CoV N protein (90% similarity to SARS-CoV-2) were able to neutralize SARS-CoV-2 by PRNT (plaque reduction neutralizing test). SARS-CoV-2 serum cross-reacted with SARS-CoV S and S1 proteins, and to a lower extent with MERS-CoV S protein, but not with the MERS-CoV S1 protein, consistent with an analysis of genetic similarity. No reactivity to SARS-CoV-2 antigens was observed in serum from patients with ubiquitous human CoV infections (common cold) or to non-CoV viral respiratory infections.
13.69.3 Limitations
Authors describe development of a serological ELISA based assay for the detection of neutralizing antibodies towards regions of the spike and nucleocapsid domains of the SARS-CoV-2 virus. Serum samples were obtained from PCR-confirmed COVID-19 patients. Negative control samples include a cohort of patients with confirmed recent exposure to non-CoV infections (i.e. adenovirus, bocavirus, enterovirus, influenza, RSV, CMV, EBV) as well as a cohort of patients with confirmed infections with ubiquitous human CoV infections known to cause the common cold. The study also included serum from patients with previous MERS-CoV and SARS-CoV zoonotic infections. This impressive patient cohort allowed the authors to determine the sensitivity and specificity of the development of their in-house ELISA assay. Of note, seroconversion was observed as early as 13 days following COVID-19 onset but the authors were not clear how disease onset was determined.
13.69.4 Significance
Validated serological tests are urgently needed to map the full spread of SARS-CoV-2 in the population and to determine the kinetics of the antibody response to SARS-CoV-2. Furthermore, clinical trials are ongoing using plasma from patients who have recovered from SARS-CoV-2 as a therapeutic option. An assay such as the one described in this study could be used to screen for strong antibody responses in recovered patients. Furthermore, the assay could be used to screen health care workers for antibody responses to SARS-CoV-2 as personal protective equipment continues to dwindle. The challenge going forward will be to standardize and scale-up the various in-house ELISA’s being developed in independent laboratories across the world.
13.70 A brief review of antiviral drugs evaluated in registered clinical trials for COVID-19
Summary of clinical trials registered as
of March7, 2020 from U.S, Chinese, Korean, Iranian and European
registries. Out of the 353 studies identified, 115 were selected for
data extraction. 80% of the trials were randomized with parallel
assignment and the median number of planned inclusions was 63 (IRQ,
36-120). Most frequent therapies in the trials included; 1) antiviral
drugs [lopinavir/ritonavir (n-15); umifenovir (n=9); favipiravir (n=7);
redmesivir (n=5)]; 2) anti-malaria drugs [chloroquine (n-11);
hydroxychloroquine (n=7)}; immunosuppressant drugs [methylprednisolone
(n=5)]; and stem cell therapies (n=23). Medians of the total number of
planned inclusions per trial for these therapies were also included.
Stem cells and lopunavir/ritonavir were the most frequently evaluated
candidate therapies (23 and 15 trials respectively), whereas remdesivir
was only tested in 5 trials but these trials had the highest median
number of planned inclusions per trial (400, IQR 394-453). Most of the
agents used in the different trials were chosen based on preclinical
assessments of antiviral activity against SARS CoV and MERS Cov corona
viruses.
The primary outcomes of the studies were clinical (66%); virological
(23%); radiological (8%); or immunological (3%). The trials were
classified as those that included patients with severe disease only;
trials that included patients with moderate disease; and trials that
included patients with severe or moderate disease.
13.70.3 Limitations
The trials evaluated provided incomplete information:
23% of these were phase IV trials but the bulk of the trials (54%) did
not describe the phase of the study. Only 52% of the trials (n=60)
reported treatment dose and only 34% (n=39) reported the duration. A lot
of the trials included a small number of patients and the trials are
still ongoing, therefore no insight was provided on the outcome of the
trials.
13.70.4 Significance
Nonetheless, this review serves as framework for
identifying COVID-19 related trials, which can be expanded upon as new
trials begin at an accelerated rate as the disease spreads around the
world.
13.70.5 Credit
This review was undertaken by K Alexandropoulos as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.71 ACE-2 Expression in the Small Airway Epithelia of Smokers and COPD Patients: Implications for COVID-19
In bronchial epithelial samples from 3 different cohorts of
individuals, ACE-2 gene expression was found to be significantly
increased in both COPD patients and smokers relative to healthy
controls. Across all test subjects, ACE-2 gene expression was also
highly correlated with decreased forced expiratory volume in 1 second
(FEV1), which may explain the increased COVID-19 disease severity in
COPD patients. Former smokers were also found to show decreased ACE2
expression relative to current smokers and had no significant difference
when compared to non-smokers.
13.71.3 Limitations
While the upregulation of ACE-2 is an interesting
hypothesis for COVID-19 disease severity in COPD patients, this study
leaves many more unanswered questions than it addresses. Further studies
are required to show whether the specific cell type isolated in these
studies is relevant to the pathophysiology of COVID-19. Furthermore,
there is no attempt to show whether that increased ACE-2 expression
contributes to greater disease severity. Does the increased ACE-2
expression lead to greater infectivity with SARS-CoV-2? There is no
mechanistic explanation for why ACE-2 levels are increased in COPD
patients. The authors could also have considered the impact of
co-morbidities and interventions such as corticosteroids or
bronchodilators on ACE-2 expression. Finally, given the extensive
sequencing performed, the authors could have conducted significantly
more in-depth analyses into gene signature differences.
13.71.4 Significance
This study attempts to address an
important clinical finding that both smokers and COPD patients show
increased mortality from COVID-19. The novel finding that ACE-2
expression is induced in smokers and COPD patients suggests not only a
mechanism for the clinical observation, but also highlights the
potential benefit of smoking cessation in reducing the risk of severe
COVID-19 disease.
13.71.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.72 Dynamic profile of severe or critical COVID-19 cases
Authors evaluate clinical correlates of 10 patients (6 male
and 4 female) hospitalized for severe Acute Respiratory Syndrome
Coronavirus 2 (SARS-CoV-2). All patients required oxygen support and
received broad spectrum antibiotics and 6 patients received anti-viral
drugs. Additionally, 40% of patients were co-infected with influenza A.
All 10 patients developed lymphopenia, two of which developed
progressive lymphopenia (PLD) and died. Peripheral blood (PB)
lymphocytes were analyzed – low CD4 and CD8 counts were noted in most
patients, though CD4:CD8 ratio remained normal.
13.72.3 Limitations
The authors evaluated a small cohort of severe
SARS-CoV-2 cases and found an association between T cell lymphopenia and
adverse outcomes. However, this is an extremely small and diverse cohort
(40% of patients were co-infected with influenza A). These findings need
to be validated in a larger cohort. Additionally, the value of this data
would be greatly increased by adding individual data points for each
patient as well as by adding error bars to each of the figures.
13.72.4 Significance
This study provides a collection of clinical data and
tracks evolution of T lymphocyte in 10 patients hospitalized for
SARS-CoV-2, of which 4 patients were co-infected with influenza A.
13.72.5 Credit
This review was undertaken by Katherine Lindblad and Tamar Plitt as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.73 Association between Clinical, Laboratory and CT Characteristics and RT-PCR Results in the Follow-up of COVID-19 patients
Data analyzed from 52 COVID-19 patients admitted
and then discharged with COVID-19. Clinical, laboratory, and
radiological data were longitudinally recorded with illness timecourse
(PCR + to PCR-) and 7 patients (13.5%) were readmitted with a follow up
positive test (PCR+) within two weeks of discharge.
13.73.3 Main Findings
At admission:
The majority of patients had increased CRP at admission (63.5%).
LDH, and HSST TNT were significantly increased at admission.
Radiographic signs via chest CT showed increased involvement in
lower lobes: right lower lobe (47 cases, 90.4%), left lower lobe
(37 cases, 71.2%).
GGO (90.4%), interlobular septal thickening (42.3%), vascular
enlargement (42.3%), and reticulation (11.5%) were most commonly
observed.
After negative PCR test (discharge):
CRP levels decreased lymphocyte counts (#/L) increased
significantly (CD3+, CD3+/8+ and CD3+/4+) after negative PCR.
Consolidation and mixed GGO observed in longitudinal CT imaging
w different extents of inflammatory exudation in lungs, with
overall tendency for improvement (except 2/7 patients that were
readmitted after discharge with re-positive test) after negative
PCR.
Seven patients repeated positive RT-PCR test and were readmitted to
the hospital (9 to 17 day after initial discharge).
Follow up CT necessary to monitor improvement during recovery
and patients with lesion progression should be given more
attention.
Dynamic CT in addition to negative test essential in clinical
diagnosis due to nasal swab PCR sampling bias (false-negatives).
Increase in CRP occurred in 2 readmitted patients (and decr. in
lymphocytes in one patient), but was not correlated with new
lesions or disease progression vs. improvement (very low N).
Patients readmitted attributed to false-negative PCR vs.
re-exposure.
13.73.4 Limitations
Patients sampled in this study were generally
younger (65.4% < 50 yrs) and less critically ill/all discharged.
Small number of recovered patients (N=18). Time of follow up was
relatively short.. Limited clinical information available about patients
with re-positive test (except CRP and lymph tracking).
13.73.5 Extended Results
NOTE: Patients sampled in this study were generally younger (65.4%
< 50 yrs) and less critically ill/all discharged. After two
consecutive negative PCR tests, patients were discharged.
Clinical Results at Admission
Median interval disease onset to admission (5 days, IQR: 3-7)
Most common symptoms included fever, fatigue, dry cough, and
expectoration.
Fifteen patients had reduced lymphocyte counts (28.8%).
No change in WBC or Neutrophil counts.
The majority of patients had increased CRP at admission (63.5%).
LDH, and HSST TNT were significantly increased at admission.
Fibrinogen was trending high though not significant.
No major changes in liver function observed.
Radiographic signs via chest CT showed increased involvement in
lower lobes: right lower lobe (47 cases, 90.4%), left lower lobe (37
cases, 71.2%).
GGO (90.4%), interlobular septal thickening (42.3%), vascular
enlargement (42.3%), and reticulation (11.5%) were most commonly
observed.
Change in Clinical Results following Negative Test
CRP levels decreased after negative PCR.
Lymphocyte counts (#/L) increased significantly (CD3+, CD3+/8+
and CD3+/4+).
No significant change to CD4/8 ratio.
LDH, HSST TNT, and Fibronegin remained high throughout, though range
observed decreased over time.
Consolidation and mixed GGO observed in longitudinal CT imaging.
Patients showed different extents of inflammatory exudation in
lungs, with overall tendency for improvement (except 2/7 patients
that were readmitted after discharge with re-positive test).
Patients Readmitted with PCR+ test
Seven patients repeated positive RT-PCR test and were readmitted
to the hospital (9 to 17 day after initial discharge).
Improvement during readmission in 4 patients and observation of
segmental progression CT in 2 patients (2/18 or 11.1% - re-positive
9 and 10 days post-discharge).
Two patients showed new GGO, while others improved greatly.
Follow up CT necessary to monitor improvement during recovery and
patients with lesion progression should be given more attention.
Dynamic CT in addition to negative test essential in clinical
diagnosis due to nasal swab PCR sampling bias (false-negatives).
Increase in CRP occurred in 2 readmitted patients (and decr. in
lymphocytes in one patient), but was not correlated with new lesions
or disease progression vs. improvement (very low N).
13.73.6 Significance
Study tracked key clinical features associated with
disease progression, recovery, and determinants of clinical
diagnosis/management of COVID-19 patients.
13.73.7 Credit
This review was undertaken by Natalie Vaninov as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.74 An orally bioavailable broad-spectrum antiviral inhibits SARS-CoV-2 and multiple 2 endemic, epidemic and bat coronavirus
ribonucleoside analog β-D-N4 30 hydroxycytidine (NHC)
Remdesivir
13.74.2 Main Findings
β-D-N4 30 –hydroxycytidine (NHC, EIDD-1931) is an
orally bioavailable ribonucleoside with antiviral activity against
various RNA viruses including Ebola, Influenza and CoV. NHC activity
introduceds mutations in the viral (but not cellular) RNA in a
dose dependent manner that directly correlated with a decrease in
viral titers. Authors show that NHC inhibited multiple genetically
distinct Bat-CoV viruses in human primary epithelial cells without
affectingcell viability even at high concentrations (100µM).
Prophylactic oral administration of NHC in C57BL/6 mice reduce lung
titers of SARS-CoV and prevented weight loss and hemorrhage. Therapeutic
administration of NHC in C57BL/6 mice 12 hours post infected with
SARS-CoV reduced acute lung injury, viral titer, and lung hemorrhage.
The degree of clinical benefit was dependent on the time of treatment
initiation post infection. The authors also demonstrate that NHC reduces
MERS-CoV infection titers, pathogenesis, and viral RNA in prophylactic
and therapeutic settings.
13.74.3 Limitations
Most of the experiments were conducted using MERS-CoV, and
SARS-CoV and a few experiments were conducted using other strains of CoV
as opposed to SARS-CoV-2. The authors note the core residues that make
up the RNA interaction sites (which constitutes the NHC interaction
sites) are highly conserved among CoV and because of this conservation
their understanding is that NHC can inhibit a broad-spectrum of CoV
including SARS-CoV-2.
The increased viral mutation rates associated with NHC activity may have
adverse effects if mutations cause the virus to become drug resistant,
more infectious or speed-up immune evasion. In addition, the temporal
diminishing effectiveness of NHC on clinical outcome when NHC was used
therapeutically is concerning. However, the longer window (7-10 days)
for clinical disease onset in human patients from the time of infection
compared to that of mice (24-48 hours), may associate with increased NHC
effectiveness in the clinic.
13.74.4 Significance
Prophylactic or therapeutic oral administration of NHC
reduces lung titers and prevents acute lung failure in C57B\6 mice
infected with CoV. Given its broad-spectrum antiviral activity, NHC
could turn out to be a useful drug for treating current, emerging and
future corona virus outbreaks.
#### Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.75 Identification of antiviral drug candidates against SARS-CoV-2 from FDA-approved drugs
A panel of ~3,000 FDA- and IND-approved
antiviral drugs were previously screened for inhibitory efficacy against
SARS CoV, a coronavirus related to the novel coronavirus SARS CoV-2
(79.5%) homology. 35 of these drugs along with another 15 (suggested by
infectious disease specialists) were tested in vitro for their ability
to inhibit SARS CoV-2 infectivity of Vero cells while preserving cell
viability. The infected cells were scored by immunofluorescence analysis
using an antibody against the N protein of SARS CoV-2. Chloroquine,
lopinavir and remdesivir were used as reference drugs.
Twenty four out of 50 drugs exhibited antiviral activity with IC50
values ranging from 0.1-10µM. Among these, two stood ou: 1) the-anti
helminthic drug niclosamide which exhibited potent antiviral activity
against SARS CoV-2 (IC50=0.28 µM). The broad-spectrum antiviral effect
of niclosamide against SARS and MERS-CoV have been previously documented
and recent evidence suggests that in may inhibit autophagy and reduce
MERS C0V replication. 2) Ciclesonide, a corticosteroid used to treat
asthma and allergic rhinitis, also exhibited antiviral efficacy but with
a lower IC50 (4.33µM) compared to niclosamide. The antiviral effects of
ciclesonide were directed against NSP15, a viral riboendonuclease which
is the molecular target of this drug.
13.75.3 Limitations
The drugs were tested against SARS CoV-2 infectivity in
vitro only, therefore preclinical studies in animals and clinical trials
in patients will be need for validation of these drugs as therapeutic
agents for COVID-19. In addition, niclosamide exhibits low adsorption
pharmatokinetically which could be alleviated with further development
of drug formulation to increase effective delivery of this drug to
target tissues. Nonetheless, niclosamide and ciclesonide represent
promising therapeutic agents against SARS CoV-2 given that other
compounds tested in the same study including favipiravir (currently used
in clinical trials) and atazanavir (predicted as the most potent
antiviral drug by AI-inference modeling) did not exhibit antiviral
activity in the current study.
13.75.4 Credit
This review was undertaken by K Alexandropoulos as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.76 Respiratory disease and virus shedding in rhesus macaques inoculated with SARS-CoV-2
animal model, pulmonary infiltrates, dynamic
of antibody response, cytokine
13.76.2 Main Findings
Inoculation of 8 Resus macaques with SARS-Cov-2 , which
all showed clinical signs of infection (respiratory pattern, reduced
appetite, weight loss, elevated body temperature) resulting in moderate,
transient disease. Four animals were euthanized at 3dpi, the 4 others at
21 dpi. Study of viral loads in different organs showed that nose swab
and throat swabs were the most sensitive, with bronchio-alveolar lavage.
Interstitial pneumonia was visible in radiographies and at the
histological scale too. Clinically, the macaques had similar symptoms as
described in human patients with moderate disease.
Viral shedding was consistently detected in nose swabs and throat swabs
immediately after infection but less consistent thereafter which could
reflect virus administration route (intranasal, oral). Bronchoalveolar
lavages performed as a measure of virus replication in the lower
respiratory tract on animals maintained for 21 days, contained high
viral loads in 1 and 3dpi. The majority of the animals exhibited
pulmonary edema and mild to moderate interstitial pneumonia on terminal
bronchioles. In addition to the lung, viral RNA could also be detected
throughout the respiratory track where viral replication mainly
occurred.
Immunologic responses included leukocytosis, neutrophilia, monocytosis
and lymphopenia in the majority of the animals at 1dpi. Lymphocytes and
monocytes re-normalized at 2dpi. Neutrophils declined after 3dpi and
through 10dpi after which they started to recover. After infection,
serum analysis revealed significant increases in IL1ra, IL6, IL10,
IL15, MCP-1, MIP-1b, but quick normalization (3dpi). Antibody
response started around 7dpi, and the antibody titers stayed elevated
until 21dpi (day of animal euthanasia).
13.76.3 Limitations
The macaques were inoculated via a combination
of intratracheal, intranasal, ocular and oral routes, which might not
reproduce how humans get infected. Maybe this can lead to different
dynamics in the host immune response. Also, the authors noted that the
seroconversion was not directly followed by a decline in viral loads, as
observed in covid19 patients.
13.76.4 Significance
This work confirms that rhesus
macaques can be a good model to study Covid-19, as it has been shown by
other groups [1315,1329,1342]. While these experiments recapitulate
moderate COVID-19 in humans, the mode of inoculation via a combination
of intratracheal, intranasal, ocular and oral routes, might not
reproduce how humans get infected and may lead to different dynamics in
the host immune response. For example, the authors noted that the
seroconversion was not directly followed by a decline in viral loads, as
observed in COVID-19 patients. Therefore, it will be interesting to
follow their antibody titers longer and further assess the
possibility/effect of reinfection in these macaques. It is essential to
be able to understand the dynamic of the disease and associated immune
responses, and to work on vaccine development and antiviral drug
testing.
13.76.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.77 ACE2 Expression is Increased in the Lungs of Patients with Comorbidities Associated with Severe COVID-19
Transcriptomic analysis using systems-level meta-analysis and
network analysis of existing literature to determine ACE2 regulation
in patients who have frequent COVID-19 comorbidities [eg-
cardiovascular diseases, familial pulmonary hypertension, cancer].
Enrichment analyses indicated pathways associated with inflammation,
metabolism, macrophage autophagy, and ER stress.
ACE2 higher in adenocarcinoma compared to adjacent normal lung; ACE2
higher in COPD patients compared to normal.
Co-expression analysis identified genes important to viral entry
such as RAB1A, ADAM10, HMGBs, and TLR3 to be associated with ACE2 in
diseased lungs.
ACE2 expression could be potentially regulated by enzymes that
modify histones, including HAT1, HDAC2, and KDM5B.
13.77.3 Limitations:
Not actual CoVID-19 patients with co-morbidities, so interpretations
in this study need to be confirmed by analyzing upcoming
transcriptomics from CoVID-19 patients having co-morbidity metadata.
As mentioned by authors, study does not look at diabetes and
autoimmunity as risk factors in CoVID-19 patients due to lack of
data; would be useful to extend such analyses to those datasets when
available.
Co-expression analysis is perfunctory and needs
validation-experiments especially in CoVID-19 lung samples to mean
anything.
Epigenomic analyses are intriguing but incomplete, as existence of
histone marks does not necessarily mean occupancy. Would be
pertinent to check cell-line data (CCLE) or actual CoVID-19 patient
samples to confirm ACE2 epigenetic control.
13.77.4 Significance
Study implies vulnerable populations have ACE2 upregulation that
could promote CoVID-19 severity. Shows important data-mining
strategy to find gene-networks associated with ACE2 upregulation in
co-morbid patients.
Several of the genes co-upregulated with ACE2 in diseased lung might
play an important role in CoVID-19 and can be preliminary targets
for therapeutics
If in silico findings hold true, epigenetic control of ACE2
expression could be a new target for CoVID-19 therapy with
strategies such as KDM5 demethylases.
13.77.5 Credit
Review by Samarth Hegde as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
This work is based on previous work by the same group
that demonstrated that SARS-CoV-2can also enter host cells via CD147
(also called Basigin, part of the immunoglobulin superfamily, is
expressed by many cell types) consistent with their previous work with
SARS-CoV-1. 1 A prospective clinical trial was conducted with 17
patients receiving Meplazumab, a humanized anti-CD147 antibody, in
addition to all other treatments. 11 patients were included as a control
group (non-randomized).
They observed a faster overall improvement rate in the Meplazumab group
(e.g. at day 14 47% vs 17% improvement rate) compared to the control
patients. Also, virological clearance was more rapid with median of 3
days in the Meplazumab group vs 13 days in control group. In laboratory
values, a faster normalization of lymphocyte counts in the Meplazumab
group was observed, but no clear difference was observed for CRP levels.
13.78.3 Limitations
While the results from the study are encouraging, this
study was non-randomized, open-label and on a small number of patients,
all from the same hospital. It offers evidence to perform a larger scale
study. Selection bias as well as differences between treatment groups
(e.g. age 51yo vs 64yo) may have contributed to results. The authors
mention that there was no toxic effect to Meplazumab injection but more
patient and longer-term studies are necessary to assess this.
13.78.4 Significance
These results seem promising as for now there are
limited treatments for Covid-19 patients, but a larger cohort of patient
is needed. CD147 has already been described to facilitate HIV [1345],
measles virus [1346], and malaria [1347] entry into host cells. This group was
the first to describe the CD147-spike route of SARS-Cov-2 entry in host
cells [1318] p147. Indeed, they had previously shown in 2005 that SARS-Cov
could enter host cells via this transmembrane protein [1348]. Further
biological understanding of how SARS-CoV-2 can enter host cells and how
this integrates with ACE2R route of entry is needed. Also, the specific
cellular targets of the anti-CD147 antibody need to be assessed, as this
protein can be expressed by many cell types and has been shown to
involved in leukocytes aggregation [1349]. Lastly, Meplazumab is not a
commercially-available drug and requires significant health resources to
generate and administer which might prevent rapid development and use.
13.78.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.79 Potent human neutralizing antibodies elicited 1 by SARS-CoV-2 infection
In this study the authors report the affinity, cross reactivity (with
SARS-CoV and MERS-CoV virus) and viral neutralization capacity of 206
monoclonal antibodies engineered from isolated IgG memory B cells of
patients suffering from SARS-CoV-2 infection in Wuhan, China. All
patients but one recovered from disease. Interestingly, the patient that
did not recover had less SARS-CoV-2 specific B cells circulating
compared to other patients.
Plasma from all patients reacted to trimeric Spike proteins from
SARS-CoV-2, SARS-CoV and MERS-CoV but no HIV BG505 trimer. Furthermore,
plasma from patients recognized the receptor binding domain (RBD) from
SARS-CoV-2 but had little to no cross-reactivity against the RBD of
related viruses SARS-CoV and MERS-CoV, suggesting significant
differences between the RBDs of the different viruses. Negligible levels
of cross-neutralization using pseudoviruses bearing Spike proteins of
SARS-CoV-2, SARS-CoV or MERS-CoV, were observed, corroborating the ELISA
cross-reactivity assays on the RBDs.
SARS-CoV-2 RBD specific B cells constituted 0.005-0.065% of the total B
cell population and 0.023-0.329% of the memory subpopulation. SARS-CoV
specific IgG memory B cells were single cell sorted to sequence the
antibody genes that were subsequently expressed as recombinant IgG1
antibodies. From this library, 206 antibodies with different binding
capacities were obtained. No discernible patterns of VH usage were found
in the 206 antibodies suggesting immunologically distinct responses to
the infection. Nevertheless, most high-binding antibodies were derived
by clonal expansion. Further analyses in one of the patient derived
clones, showed that the antibodies from three different timepoints did
not group together in phylogenetic analysis, suggesting selection during
early infection.
Using surface plasmon resonance (SPR) 13 antibodies were found to have
10-8 tp 10-9 dissociation constants (Kd). Of the 13 antibodies, two
showed 98-99% blocking of SARS-CoV-2 RBD-ACE2 receptor binding in
competition assays. Thus, low Kd values alone did not predict ACE2
competing capacities. Consistent with competition assays the two
antibodies that show high ACE2 blocking (P2C-2F6 and P2C-1F11) were the
most capable of neutralizing pseudoviruses bearing SARS-CoV-2 spike
protein (IC50 of 0.06 and 0.03 µg/mL, respectively). Finally, using
SPR the neutralizing antibodies were found to recognize both overlapping
and distinct epitopes of the RBD of SARS-CoV-2.
13.79.3 Limitations
Relatively low number of patients
No significant conclusion can be drawn about the possible
> correlation between humoral response and disease severity
In vitro Cytopathic Effect Assay (CPE) for neutralization activity
Huh7 cells were used in neutralization assays with
> pseudoviruses, and they may not entirely mimic what happens in
> the upper respiratory tract
CPE assay is not quantitative
Duplicated panel in Figure 4C reported (has been fixed in version 2)
13.79.4 Significance
This paper offers an explanation as to why
previously isolated antibodies against SARS-CoV do not effectively block
SARS-CoV-2. Also, it offers important insight into the development of
humoral responses at various time points during the first weeks of the
disease in small but clinically diverse group of patients. Furthermore,
it provides valuable information and well characterized antibody
candidates for the development of a recombinant antibody treatment for
SARS-CoV-2. Nevertheless, it also shows that plasmapheresis might have
variability in its effectiveness, depending on the donor’s antibody
repertoire at the time of donation.
13.79.5 Credit
Review by Jovani Catalan-Dibene as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.80 Characterisation of the transcriptome and proteome of SARS-CoV-2 using direct RNA sequencing and tandem mass spectrometry reveals evidence for a cell passage induced in-frame deletion in the spike glycoprotein that removes the furin-like cleavage site.
The authors performed long read RNA sequencing using an Oxford Nanopore
MinION as well as tandem mass spec (MS) on Vero cells (a cell line
derived from kidney cells of the African green monkey that is deficient
in interferon) infected with SARS-CoV-2.
The authors found that passage of the virus in Vero cells gave rise to a
spontaneous 9 amino acid deletion (679-NSPRRARSV-687 to I) in the spike
(S) protein. The deleted sequence overlaps a predicted furin cleavage
site at the S1 / S2 domain boundary that is present in SARS-CoV-2 but
not SARS-CoV or the closely related bat coronavirus RaTG13, which are
cleaved at S1 / S2 by other proteases [54]. Furin cleavage sites at
similar positions in other viruses have been linked to increased
pathogenicity and greater cell tropism [1351]. Loss of this site in
SARS-CoV-2 has also already been shown to increase viral entry into Vero
but not BHK cells (which are also interferon deficient) [67]. The
authors therefore make an important contribution in demonstrating that
passage in Vero cells may lead to spontaneous loss of a key
pathogenicity-conferring element in SARS-CoV-2.
13.80.3 Limitations
As the authors note, a similar study posted earlier by Kim et al., which
also passaged SARS-CoV-2 in Vero cells, did not identify any loss in the
S protein furin cleavage site [1352]. It therefore remains to be
determined how likely it is that this mutation spontaneously arises. A
quantitative investigation using multiple experimental replicas to
understand the spontaneous viral mutation rate at this site and
elsewhere would be informative. Also, the mechanistic basis for the
higher viral fitness conferred by loss of the furin cleavage site in
Vero cells – but, evidently, not in vivo in humans, as this site is
maintained in all currently sequenced circulating isolates - remains to
be understood.
Due to the high base-call error rate of MinION sequencing, the authors’
bioinformatic pipeline required aligning transcripts to a reference to
correct sequencing artifacts. This presumably made it difficult or
impossible to identify other kinds of mutations, such as single
nucleotide substitutions, which may occur even more frequently than the
deletions identified in this work. Pairing long read sequencing with
higher-accuracy short-read sequencing may be one approach to overcome
this issue.
13.80.4 Significance
As the authors suggest, animal studies using live virus challenge may
need to periodically verify the genomic integrity of the virus, or
potentially risk unknowingly using a likely less-pathogenic variant of
the virus.
More broadly, the results emphasize the complexity and plasticity of the
SARS-CoV-2 viral transcriptome and proteome. For example, the authors
found multiple versions of transcripts encoding the nucleocapsid (N)
protein, each with different small internal deletions, some of which
were verified for translation by MS. A number of peptides arising from
translation of unexpected rearrangements of transcripts were also
detected. Additionally, the authors identified phosphorylation of a
number of viral proteins (N, M, ORF 3a, nsp3, nsp9, nsp12 and S). For
any cases where these have functional consequences, targeting the
kinases responsible could be an avenue for drug development.
Understanding the functional consequences of the mutations, transcript
variations, and post translational modifications identified in this
study will be important future work.
13.80.5 Credit
This review was undertaken by Tim O’Donnell, Maria Kuksin as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.81 A SARS-CoV-2-Human Protein-Protein Interaction Map Reveals Drug Targets and Potential Drug- Repurposing
Gordon et al cloned, tagged and expressed 26 of the 29
SARS-CoV-2 proteins individually in HEK293T cells and used mass
spectrometry to identify protein-protein interactions. They identified
332 viral-host protein-protein interactions. Furthermore, they used
these interactions to identify 66 existing drugs known to target host
proteins or host pathways (eg SARS-CoV-2 N and Orf8 proteins interact
with proteins regulated by the mTOR pathway, so mTOR inhibitors
Silmitasertib and Rapamycin are possible drug candidates).
13.81.3 Limitations
The main limitation of the study stems from
the reductionist model: overexpression of plasmids encoding individual
viral proteins in HEK293T cells. This precludes any interactions between
the viral proteins, or the combined effects of multiple proteins on the
host, as they are expressed individually. Moreover, HEK293T cells come
from primary embryonic kidney and therefore might not reflect how
SARS-CoV-2 interacts with its primary target, the lung. However, the
authors found that the proteins found to interact with viral proteins in
their experiments are enriched in lung tissue compared to HEK293Ts.
13.81.4 Significance
The authors provide a “SARS-CoV-2 interaction map,”
which may provide potential hypotheses as to how the virus interacts
with the host. Further, they identified existing drugs that could
disrupt these host-viral interactions and curb SARS-CoV-2 infection.
Although these interactions have not been validated, this paper acts as
a valuable resource.
13.81.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.82 First Clinical Study Using HCV Protease Inhibitor Danoprevir to Treat Naïve and Experienced COVID-19 Patients
The authors treated 11 Covid-19 patients with Danoprevir, a
commercialized HCV protease inhibitor [1354](p4), boosted by ritonavir [1355], a
CYP3A4 inhibitor (which enhances the plasma concentration and
bioavailabilty of Danoprevir). Two patients had never received
anti-viral therapy before (=naïve), whereas nine patients were on
Lopinavir/Ritonavir treatment before switching to Danoprevir/Ritonavir
(=experienced). The age ranged from 18 to 66yo.
Naïve patients that received Danoprevir/Ritonavir treatment had a
decreased hospitalization time. Patients treated with
Lopinavir/Ritonavir did not have a negative PCR test, while after
switching to Danoprevir/Ritonavir treatment, the first negative PCR test
occurred at a median of two days.
13.82.3 Limitations
The results of the study are very hard to interpret as there is no
control group not receiving Danoprevir/Ritonavir treatment. This was
especially true in naïve patients who seemed to have more mild symptoms
before the start of the study and were younger (18 and 44yo) compared to
the experienced patients (18 to 66yo). The possibility that the patients
would have recovered without Danoprevir/Ritonavir treatment cannot be
excluded.
13.82.4 Significance
The authors of the study treated patients with Danoprevir, with the
rational to that this is an approved and well tolerated drug for HCV
patients [1355], and that it could also target the protease from SARS-CoV-2
(essential for viral replication and transcription). Indeed, homology
modelling data indicated that HCV protease inhibitors have the highest
binding affinity to Sars-Cov2 protease among other approved drugs [1356].
While this study shows that the combination of Danoprevir and Ritonavir
might be beneficial for Covid-19 patients, additional clinical trials
with more patients and with better methodology (randomization and
control group) are needed to make further conclusions.
13.82.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.83 Efficacy of hydroxychloroquine in patients with COVID-19: results of a randomized clinical trial
This is a randomized clinical trial of hydroxychloroquine (HCQ) efficacy
in the treatment of COVID-19. From February 4 – February 28, 2020 142
COVID-19 positive patients were admitted to Renmin Hospital of Wuhan
University. 62 patients met inclusion criteria and were enrolled in a
double blind, randomized control trial, with 31 patients in each arm.
Inclusion criteria:
Age ≥ 18 years
Positive diagnosis COVID-19 by detection of SARS-CoV-2 by RT-PCR
Diagnosis of pneumonia on chest CT
Mild respiratory illness, defined by SaO2/SPO2 ratio > 93% or
PaO2/FIO2 ratio > 300 mmHg in hospital room conditions (Note:
relevant clinical references described below.)
Hypoxia is defined as an SpO2 of 85-94%; severe hypoxia <
85%.
The PaO2/FIO2 (ratio of arterial oxygen tension to fraction
of inspired oxygen) is used to classify the severity of acute
respiratory distress syndrome (ARDS). Mild ARDS has a
PaO2/FIO2 of 200-300 mmHg, moderate is 100-200, and severe
< 100.
Willing to receive a random assignment to any designated treatment
group; not participating in another study at the same time
Exclusion criteria:
Severe or critical respiratory illness (not explicitly defined,
presumed to be respiratory function worse than outlined in inclusion
criteria); or participation in trial does not meet patient’s maximum
benefit or safe follow up criteria
Retinopathy or other retinal diseases
Conduction block or other arrhythmias
Severe liver disease, defined by Child-Pugh score ≥ C or AST >
twice the upper limit
Pregnant or breastfeeding
Severe renal failure, defined by eGFR ≤ 30 mL/min/1.73m2, or on
dialysis
Potential transfer to another hospital within 72h of enrollment
Received any trial treatment for COVID-19 within 30 days before the
current study
All patients received the standard of care: oxygen therapy, antiviral
agents, antibacterial agents, and immunoglobulin, with or without
corticosteroids. Patients in the HCQ treatment group received additional
oral HCQ 400 mg/day, given as 200 mg 2x/day. HCQ was administered from
days 1-5 of the trial. The primary endpoint was 5 days post enrollment
or a severe adverse reaction to HCQ. The primary outcome evaluated was
time to clinical recovery (TTCR), defined as return to normal body
temperature and cough cessation for > 72h. Chest CT were imaged on
days 0 and 6 of the trial for both groups; body temperature and patient
reports of cough were collected 3x/day from day 0 – 6. The mean age and
sex distribution between the HCQ and control arms were comparable.
13.83.3 Main Findings
There were 2 patients showing mild secondary effects of HCQ treatment.
More importantly, while 4 patients in the control group progressed to
severe disease, none progressed in the treatment group.
TTCR was significantly decreased in the HCQ treatment arm; recovery from
fever was shortened by one day (3.2 days control vs. 2.2 days HCQ, p =
0.0008); time to cessation of cough was similarly reduced (3.1 days
control vs. 2.0 days HCQ, p = 0.0016).
Overall, it appears that HCQ treatment of patients with mild COVID-19
has a modest effect on clinical recovery (symptom relief on average 1
day earlier) but may be more potent in reducing the progression from
mild to severe disease.
13.83.4 Limitations
This study is limited in its inclusion of only patients with mild
disease, and exclusion of those on any treatment other than the standard
of care. It would also have been important to include the laboratory
values of positive RT-PCR detection of SARS-CoV-2 to compare the
baseline and evolution of the patients’ viral load.
13.83.5 Limitations
Despite its limitations, the study design has good rigor as a double
blind RCT and consistent symptom checks on each day of the trail. Now
that the FDA has approved HCQ for treatment of COVID-19 in the USA, this
study supports the efficacy of HCQ use early in treatment of patients
showing mild symptoms, to improve time to clinical recovery, and
possibly reduce disease progression. However, most of the current
applications of HCQ have been in patients with severe disease and for
compassionate use, which are out of the scope of the findings presented
in this trial. Several additional clinical trials to examine
hydroxychloroquine
are now undergoing; their results will be critical to further validate
these findings.
13.83.6 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
Structure-based modeling of SARS-CoV-2 peptide/HLA-A02 antigens
The authors utilize homology modeling to identify peptides from the
SARS-CoV-2 proteome that potentially bind HLA-A*02:01.
They utilize high-resolution X-ray structures of peptide/MHC
complexes on Protein Data Bank, substitute homologous peptides with
SARS-CoV-2 peptides, and calculate MHC/SARS-CoV-2 peptide Rosetta
binding energy.
They select MHC/SARS-CoV-2 complex models with highest binding
energy for further study and publish models in an online database
(https://rosettamhc.chemistry.ucsc.edu).
Limitations:
The authors only utilize computational methods and predicted
SARS-CoV-2 peptides must be validated experimentally for
immunogenicity and clinical response.
Due to computational burden and limited availability of high
resolution X-ray structures on PDB, authors only simulate 9-mer and
10-mer peptide binding to HLA-A*02:01.
Since the authors compare select existing X-ray structures as a
starting point, backbone conformations that deviate significantly
between test and template peptides are not captured. Furthermore,
Rosetta modeling protocols do not capture all possible structures
and binding energy scoring does not fully recapitulate fundamental
forces.1,2
Importance/Relevance:
The authors identify and publish high-scoring SARS-CoV-2 peptides
that may direct a targeted, experimental validation approach toward
a COVID-19 vaccine.
The authors utilize Rosetta simulation to further filter results
from NetMHCpan 4.0, supporting machine learning prediction with
structural analysis.
The authors develop RosettaMHC, a computationally efficient method
of leveraging existing X-ray structures for identification of
immunogenic peptides.
References:
Bender, B. J., Cisneros, A., 3rd, Duran, A. M., Finn, J. A., Fu, D.,
Lokits, A. D., . . . Moretti, R. (2016). Protocols for Molecular
Modeling with Rosetta3 and RosettaScripts. Biochemistry, 55(34),
4748-4763. doi:10.1021/acs.biochem.6b00444
Alford, R. F., Leaver-Fay, A., Jeliazkov, J. R., O’Meara, M. J.,
DiMaio, F. P., Park, H., . . . Gray, J. J. (2017). The Rosetta
All-Atom Energy Function for Macromolecular Modeling and Design. J
Chem Theory Comput, 13(6), 3031-3048. doi:10.1021/acs.jctc.7b00125
Review by Jonathan Chung as part of a project by students, postdocs and
faculty at the Immunology Institute of the Icahn school of medicine,
Mount Sinai.
13.84 Serology characteristics of SARS-CoV-2 infection since the exposure and post symptoms onset
Currently, the diagnosis of SARS-CoV-2 infection
entirely depends on the detection of viral RNA using polymerase chain
reaction (PCR) assays. False negative results are common, particularly
when the samples are collected from upper respiratory. Serological
detection may be useful as an additional testing strategy. In this study
the authors reported that a typical acute antibody response was induced
during the SARS-CoV-2 infection, which was discuss earlier1. The
seroconversion rate for Ab, IgM and IgG in COVID-19 patients was 98.8%
(79/80), 93.8% (75/80) and 93.8% (75/80), respectively. The first
detectible serology marker was total antibody followed by IgM and IgG,
with a median seroconversion time of 15, 18 and 20 days-post exposure
(d.p.e) or 9, 10- and 12-days post-onset (d.p.o). Seroconversion was
first detected at day 7d.p.e in 98.9% of the patients. Interestingly
they found that viral load declined as antibody levels increased. This
was in contrast to a previous study [1297], showing that increased antibody
titers did not always correlate with RNA clearance (low number of
patient sample).
13.84.3 Limitations
Current knowledge of the antibody response to SAR-CoV-2
infection and its mechanism is not yet well elucidated. Similar to the
RNA test, the absence of antibody titers in the early stage of illness
could not exclude the possibility of infection. A diagnostic test, which
is the aim of the authors, would not be useful at the early time points
of infection but it could be used to screen asymptomatic patients or
patients with mild disease at later times after exposure.
13.84.4 Significance
Understanding the antibody responses against SARS-CoV2 is
useful in the development of a serological test for the diagnosis of
COVID-19. This manuscript discussed acute antibody responses which can
be deducted in plasma for diagnostic as well as prognostic purposes.
Thus, patient-derived plasma with known antibody titers may be used
therapeutically for treating COVID-19 patients with severe illness.
13.84.5 Credit
This review was undertaken and edited by Konstantina A as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.85 SARS-CoV-2 launches a unique transcriptional signature from in vitro, ex vivo, and in vivo systems
Given the high mortality rate of SARS-CoV-2 relative to
other respiratory viruses such seasonal IAV and RSV, there may be
underlying host-pathogen interactions specific to SARS-CoV-2 that
predispose to a worse clinical outcome. Using in vivo, ex vivo, and
in vitro systems, the authors profiled host cell transcriptional
responses to SARS-CoV-2 and to other common respiratory viruses
(seasonal IAV and RSV). SARS-CoV-2 infection in vitro led to an
induction of type I interferon response signaling and the upregulation
of cytokine/chemokines transcripts. In comparison with IAV and RSV
infection, SARS-CoV-2 in vitro appears to uniquely induce less type I
and type III interferon expression and higher levels of two cytokines
previously implicated in respiratory inflammation. Lastly, in vivo
data from ferrets showed a reduced induction of cytokines and chemokines
by SARS-CoV-2 infection relative to IAV infection.
13.85.3 Limitations
While these results are promising, there are several
key weaknesses of this paper. 1) As the authors point out, there is an
undetectable level of SARS-CoV-2 putative receptor (ACE2) and protease
(TMPRSS2) expression in the lung epithelial cell line used for the in
vitro studies. This raises the important question of whether viral
replication actually occurs in any of the models used, which may explain
the lack of interferon production observed in vitro in SARS-CoV-2
treated cells. Further studies characterizing viral titers across
timepoints are needed. 2) Furthermore, these studies only characterize
the host response at a single dose and timepoint per virus, and it is
unclear why these doses/timepoints were chosen. This leaves open the
possibility that the observed differences between viruses could be due
to differences in dose, timing, host response, or a combination of all
of these. 3) It is unclear whether ferrets are productively infected,
which cell types are infected, and the extent/timing of the clinical
course of infection. Moreover, the in vitro and in vivo data do not
strongly correlate and the reasons for this are unclear.
13.85.4 Significance
This paper describes potentially unique
transcriptional signatures of host cells exposed to SARS-CoV-2. If
validated, these findings may help explain clinical outcomes and could
be targeted in future therapeutic interventions.
13.85.5 Potential Conflicts of Interest Disclosure
The reviewers are also researchers at the Icahn School
of Medicine at Mount Sinai.
13.85.6 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.86 A New Predictor of Disease Severity in Patients with COVID-19 in Wuhan, China
377 hospitalized patients were divided into two groups: severe and
non-severe pneumonia. The laboratory results of their first day of
admission were retrospectively analyzed to identify predictors of
disease severity.
After adjusting for confounding factors from chronic comorbidities (such
as high blood pressure, type 2 diabetes, coronary heart disease, and
chronic obstructive pulmonary disease), the independent risk factors
identified for severe pneumonia were age, the ratio of
neutrophil/lymphocytes counts, CRP and D-dimer levels.
To further increase the specificity and sensibility of these markers,
they showed that their multiplication [(Neutrophil/lymphocyte count)
* CRP * D-dimer] was a better predictor of disease severity, with
higher sensitivity (95.7%) and specificity (63.3%), with a cutoff value
of 2.68.
13.86.3 Limitations
This study included 377 hospitalized patients. Among them, 45.6%
patients tested positive for SARS-Cov-2 nucleic acid test results, and
others were included in the study based on clinically diagnosis even if
the molecular diagnosis was negative. Thus, additional studies are
needed to verify this on a larger number of covid-19 certified patients
and the cutoff value might be adjusted. Also, all the patients that did
not have the clinical characteristics of severe pneumonia were included
in the non-severe pneumonia group, but usually patients are also divided
into moderate and mild disease.
Also, studying different subset of lymphocytes could lead to a more
specific predictor. Another study showed that the neutrophils to CD8+ T
cells ratio was a strong predictor of disease severity [1268]. Another more
precise study showed that the percentage of helper T cells and
regulatory T cells decrease but the percentage of naïve helper T cells
increases in severe cases [1261]. Taking these subpopulations into account
might make the predictor more powerful.
Other studies also noted an inverse correlation between disease severity
and LDH [1301] or IL6 [1310] levels, but the authors here do not discuss LDH nor
IL6 levels, although this could help to strengthen the predictor.
The study is based on the results obtained on the first day of
admission, studying the dynamic of the changes in patients might also be
interesting to better predict disease severity.
13.86.4 Significance
This study confirms that the neutrophil to lymphocyte ratio can be a
predictor of disease severity as shown by many others [1260,1261,1274]. The
novelty here is that they show that a combination with other markers can
enhance the specificity and sensibility of the predictor, although the
study could be improved by taking into account sub-populations of
lymphocytes and more biological factors from patients such as LDH and
IL6.
13.86.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.87 Metabolic disturbances and inflammatory dysfunction predict severity of coronavirus disease 2019 (COVID-19): a retrospective study
Retrospective Study on 97 COVID-19 hospitalized patients
(25 severe and 72 non-severe) analyzing clinical and laboratory
parameter to predict transition from mild to severe disease based on
more accessible indicators (such as fasting blood glucose, serum protein
or blood lipid) than inflammatory indicators. In accordance with other
studies, age and hypertension were risk factors for disease severity,
and lymphopenia and increased IL-6 was observed in severe patients. The
authors show that fasting blood glucose (FBG) was altered and patients
with severe disease were often hyperglycemic. Data presented support
that hypoproteinaemia, hypoalbuminemia, and reduction in
high-densitylipoprotein (HDL-C) and ApoA1 were associated with disease
severity.
13.87.3 Limitations
In this study non-severe patients were divided in two groups based on
average course of the disease: mild group1 (14 days, n=28) and mild
group 2 (30 days, n=44). However mild patients with a longer disease
course did not show an intermediate phenotype (between mild patients
with shorter disease course and severe patients), hence it is unclear
whether this was a useful and how it impacted the analysis. Furthermore,
the non-exclusion of co-morbidity factors in the analysis may bias the
results (e.g. diabetic patients and glucose tests) It is not clear at
what point in time the laboratory parameters are sampled. In table 3, it
would have been interesting to explore a multivariate multiple
regression. The correlation lacks of positive control to assess the
specificity of the correlation to the disease vs. correlation in any
inflammatory case. The dynamic study assessing the predictability of the
laboratory parameter is limited to 2 patients. Hence there are several
associations with disease severity, but larger studies are necessary to
test the independent predictive value of these potential biomarkers.
13.87.4 Significance
As hospital are getting overwhelmed a set of
easily accessible laboratory indicators (such as serum total protein)
would potentially provide a triage methodology between potentially
severe cases and mild ones. This paper also opens the question regarding
metabolic deregulation and COVID-19 severity.
13.87.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.88 Viral Kinetics and Antibody Responses in Patients with COVID-19
Prospective cohort of 67 patients, clinical specimens taken and
follow-up conducted.
Viral shedding, serum IgM, IgG antibody against NP evaluated and
correlated to disease severity and clinical outcome
Viral RNA levels peaked at 1 week from febrile/cough symptom onset
in sputum, nasal swabs, and stool samples. Shedding ranged from
12-19 days (median ranges) and was longer in severe patients.
IgM and IgG titers stratified patients into three archetypes as
‘strong vs weak vs non-responders’. Strong responders (with higher
IgM/IgG titers) were significantly higher in severe patients.
13.88.3 Limitations
Specific for immune monitoring.
Not clear if stool RNA captured from live infection in
intestine/liver or from swallowed sputum. Transmission electron
microscopy (TEM) carried out on sputum samples as proof of concept,
but not stools. TEM unreasonable for actual clinical diagnosis.
Several patients had co-morbidities (such as pulmonary and liver
disease) that were not accounted for when tracking antibody
responses. Viral kinetics and IgM/IgG titers in subsets of patients
with underlying conditions/undergoing certain medication would be
informative.
13.88.4 Significance
Three archetypes of antibody response to SARS-CoV-2 with different
disease progression and kinetics is useful to stratify patients, and
for future serological tests.
Strong spike-IgG levels often correlate with lymphopenia and
CoVID-19 disease severity [1362], similar to macaque
studies in SARS [1363]. It would be critical to see if anti-NP or
anti-Spike IgG antibodies for SARS-CoV-2 also elicit similar
detrimental effects before clinical use.
13.88.5 Credit
Review by Samarth Hegde as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.89 COVID-19 infection induces readily detectable morphological and inflammation-related phenotypic changes in peripheral blood monocytes, the severity of which correlate with patient outcome
This study is based on flow cytometry immunophenotyping of
PBMCs from 28 patients diagnosed positive for SARS-Cov2 (COVID19). The
authors identify a population of abnormally large (FSC-hi) monocytes,
present in COVID19 patients, but absent in PBMCs of healthy volunteers
(n=16) or patients with different infections (AIDS, malaria, TB). This
FSC-hi monocytic population contains classical, intermediate and
non-classical (monocytes (based on CD14 and CD16 expression) that
produce inflammatory cytokines (IL-6, TNF and IL-10). The authors
suggest an association of FSC-hi monocytes with poor outcome and
correlate a high percentage of FSC-low monocytes, or higher ratio of
FSC-low/hi monocytes, with faster hospital discharge.
13.89.3 Limitations
While identification of the monocytic population based
on FSC is rather robust, the characterization of these cells remains
weak. A comprehensive comparison of FSC-hi monocytes with FSC-low
monocytes from patients and healthy controls would be of high value. It
is unclear if percentages in blood are among CD45+ cells. Furthermore,
it would have been important to include absolute numbers of different
monocytic populations (in table 1 there are not enough samples and it is
unclear what the authors show).
The authors show expression of the ACE2 receptor on the surface of the
monocytes, and highlight these cells as potential targets of SARS-Cov2.
However, appropriate controls are needed. CD16 has high affinity to
rabbit IgG and it is unclear whether the authors considered unspecific
binding of rabbit anti-ACE2 to Fc receptors. Gene expression of ACE-2 on
monocytes needs to be assessed. Furthermore, it would be important to
confirm infection of monocytes by presence of viral proteins or viral
particles by microscopy.
Considering the predictive role of FSC-hi monocytes on the development
of the disease and its severity, some data expected at this level are
neither present nor addressed. Although the cohort is small, it does
include 3 ICU patients. What about their ratio of FSC-low vs FSC-hi
monocytes in comparison to other patients? Was this apparent early in
the disease course? Does this population of FSC-hi monocytes differ
between ICU patients and others in terms of frequency, phenotype or
cytokine secretion?
In general, figures need to revised to make the data clear. For example,
in Fig. 5, according to the legend it seems that patients with FSC-high
monocytes are discharged faster from the hospital. However according to
description in the text, patients were grouped in high or low levels of
FSC-low monocytes.
13.89.4 Significance
Despite the limitations of this study, the discovery of
a FSC-high monocyte population in COVID-19 patients is of great
interest. With similar implication, a the recent study by Zhou et al. [1266]
identified a connection between an inflammatory CD14+CD16+ monocyte
population and pulmonary immunopathology leading to deleterious clinical
manifestations and even acute mortality after SARS-CoV-2 infections.
Although the presence of these monocytes in the lungs has yet to be
demonstrated, such results support the importance of monocytes in the
critical inflammation observed in some COVID19 patients.
13.89.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.90 Correlation between universal BCG vaccination policy and reduced morbidity and mortality for COVID-19: an epidemiological study
The authors compared middle and high income countries that never had a
universal BCG vaccination policy (Italy, Lebanon, Nederland, Belgium)
and countries with a current policy (low income countries were excluded
from the analysis as their number of cases and deaths might be
underreported for the moment). Countries that never implement BCG
vaccination have a higher mortality rate than countries which have a BCG
vaccination policy (16.38 deaths per million people vs 0.78). Next,
the authors show that an earlier start of vaccination correlates with
a lower number of deaths per million inhabitants. They interpret this
as the vaccine protecting a larger fraction of elderly people, which are
usually more affected by COVID-19. Moreover, higher number of COVID-19
cases were presented in countries that never implemented a universal
BCG vaccination policy.
13.90.3 Limitations
While this study aims to test an intriguing hypothesis unfortunately,
the data is not sufficient at this time to accurately make any
determinations. Several caveats must be noted including: not all
countries are in the same stage of the pandemic, the number of
cases/deaths is still changing very rapidly in a lot of countries and
thus the association may only reflect exposure to the virus. This
analysis would need to be re-evaluated when all the countries are passed
the pandemic and more accurate numbers are available. Additionally, very
few middle and high-income countries ever implemented universal BCG
vaccination, which can be a source of bias (5 countries, vs 55 that have
a BCG vaccine policy). Effective screening and social isolation policies
also varied considerable across the countries tested and may reflect
another important confounder. The authors could consider analyzing the
Case Fatality Rate (CFR, % of patients with COVID-19 that die), to more
correct for exposure although testing availability will still bias this
result. Variability in mortality within countries or cities with
variable vaccination and similar exposure could also be appropriate
although confounders will still be present.
13.90.4 Significance
BCG vaccine is a live attenuated strain derived from Mycobacterium
bovis and used for a vaccine for tuberculosis (TB). This vaccine has
been proven to be efficient in preventing childhood meningitis TB, but
doesn’t prevent adult TB as efficiently. For this reason, several
countries are now only recommending this vaccine for at-risk population
only.
This study shows that there is a correlation between BCG vaccination
policy and reduced mortality for Covid-19. Indeed, BCG vaccine has
been shown to protect against several viruses and enhance innate
immunity [1366], which could explain why it could protect against SARS-CoV-2
infection, but the exact mechanism is still unknown. Moreover, the
efficiency of adult/older people vaccination and protection against
Covid-19 still needs to be assessed. Regarding this, Australian
researchers are starting a clinical trial of BCG vaccine for healthcare
workers [1367], to assess if it can protect them against Covid-19.
13.90.5 Credit
This review was undertaken as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.91 Non-neural expression of SARS-CoV-2 entry genes in the olfactory epithelium
Study analyzed bulk and scRNAseq data of olfactory cell types from
publicly-available mouse, nonhuman primate and human datasets.
show that ACE2 and TMPRSS2 (genes involved in SARS-CoV-2 entry)
are expressed in olfactory epithelial (OE) cells, basal stem cells
and respiratory epithelium (RE), but not sensory neurons.
Comparison of human RE and OE datasets (Deprez et al. 2019; Durante
et al. 2020) revealed that ACE2 and TMPRSS2 expression in OE
sustentacular cells was similar to expression in the remainder of
the non-nasal respiratory tract.
13.91.3 Limitations
Transcript data alone from healthy respiratory/olfactory cells is
not sufficient to confirm infectivity of nasal passage, or to
indicate damage to epithelia.
No mechanism defined for anosmia; it is not clear if epithelial
injury leads to reduced sensitivity or increased inflammation and
altered immune contexture drives neural/epithelial dysfunction. Will
be critical to test this in CoVID-19 patient samples or mouse
models.
13.91.4 Significance
Study provides possible rationale for anosmia observed in several
CoVID-19 patients.
Raises possibility that nasal respiratory goblet, ciliated cells,
and olfactory epithelia may serve as a viral reservoir after initial
SARS-CoV-2 infection.
Human olfactory sensory neurons express several other molecules
important to CoV (not CoV-19) entry such as FURIN, ST6GAL1,
ST3GAL4; this suggests wider mechanism of neuronal infectivity in
other coronaviruses.
13.91.5 Credit
Review by Samarth Hegde as part of a project by students, postdocs and
faculty at the Immunology Institute of the Icahn School of Medicine,
Mount Sinai.
Title:
SARS-CoV-2 proteome microarray for mapping COVID-19 antibody
interactions at amino acid resolution
This study screened the viral protein epitopes recognized by antibodies
in the serum of 10 COVID-19 patients using a new SARS-CoV-2 proteome
peptide microarray. The peptide library was constructed with 966 linear
peptides, each 15 amino acids long with a 5 amino acid overlap, based on
the protein sequences encoded by the genome of the Wuhan-Hu-1 strain.
To investigate crossreactivity between SARS-CoV-1 and SARS-CoV-2, they
tested rabbit monoclonal and polyclonal antibodies against SARS-CoV-1
nucleocapsid (N) in the microarray. Antibodies against SARS-CoV-1 N
displayed binding to the SARS-CoV-2 nucleocapsid (N) peptides.
Polyclonal antibodies showed some crossreactivity to other epitopes from
membrane (M), spike (S), ORF1ab and ORF8. This suggests that previous
exposure to SARS-CoV-1 may induced antibodies recognizing both viruses.
Screening of IgM and IgG antibodies from 10 COVID-19 patients showed
that many antibodies targeted peptides on M, N, S, Orf1ab, Orf3a, Orf7a,
and Orf8 from SARS-CoV-2, while immunodominant epitopes with antibodies
in more than 80 % COVID-19 patients were present in N, S and Orf3. It is
shown that the receptor binding domain (RBD) resides on S protein and
RBD is important for SARS-CoV-2 to enter the host cells via ACE2. Among
six epitopes on S protein, structural analysis predicted that three
epitopes were located at the surface and three epitopes were located
inside of the protein. Furthermore, some IgM antibodies from 1 patient
and IgG antibodies from 2 patients bound to the same epitope (residue
456-460, FRKSN) which resided within the RBD, and structural analysis
determined that this epitope was located in the region of the RBD loop
that engages with ACE2.
Critical analysis of the study:
In addition to the limitations mentioned in the manuscript, it would
have been informative to do the analysis over the course of the disease.
The pattern of antibody recognition, especially on S protein, and the
course of antibodies of different isotypes recognizing the same peptide
might correlate to the clinical course in these patients. It would alos
have been informative to analyze the presence of cross-reactive
antibodies from pateints previously exposed to SARS-CoV-1.
The importance and implications for the current epidemics:
This study identified linear immunodominant epitopes on SARS-CoV-2,
Wuhan-Hu-1 strain. This is a valuable information to design vaccines
that will elicit desirable immune responses.
The Novel Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2)
Directly Decimates Human Spleens and Lymph Nodes
It has been previously reported that COVID-19 patients exhibit severe
lymphocytopenia, but the mechanism through which this depletion occurs
has not been described. In order to characterize the cause and process
of lymphocyte depletion in COVID-19 patients, the authors performed
gross anatomical and in situ immune-histochemical analyses of spleens
and lymph nodes (hilar and subscapular) obtained from post-mortem
autopsies of 6 patients with confirmed positive viremia and 3 healthy
controls (deceased due to vehicle accidents).
Primary gross observations noted significant splenic and LN atrophy,
hemorrhaging, and necrosis with congestion of interstitial blood vessels
and large accumulation of mononuclear cells and massive lymphocyte
death. They found that CD68+ CD169+ cells in the spleens, hilar and
subscapular LN, and capillaries of these secondary lymphoid organs
expressed the ACE2 receptor and stain positive for the SARS-CoV-2
nucleoprotein (NP) antigen, while CD3+ T cells and B220+ B cells
lacked both the ACE2 receptor and SARS-CoV-2 NP antigen. ACE2+ NP+
CD169+ macrophages were positioned in the splenic marginal zone (MZ)
and in the marginal sinuses of LN, which suggests that these macrophages
were positioned to encounter invading pathogens first and may contribute
to virus dissemination.
Since SARS-CoV-2 does not directly infect lymphocytes, the authors
hypothesized that the NP+ CD169+ macrophages are responsible for
persistent activation of lymphocytes via Fas::FasL interactions that
would mediate activation-induced cell death (AICD). Indeed, the
expression of Fas was significantly higher in virus-infected tissue than
that of healthy controls, and TUNEL staining showed significant
lymphocytic apoptosis. Since pro-inflammatory cytokines like IL-6 and
TNF-α can also engage cellular apoptosis and necrosis, the authors
interrogated the cytokine expression of the secondary lymphoid organs
from COVID-19 patients; IL-6, not TNF-α, was elevated in virus-infected
splenic and lymph node tissues, compared to those of healthy controls,
and immunofluorescent staining showed that IL-6 is primarily produced by
the infected macrophages. In vitro infection of THP1 cells with
SARS-CoV-2 spike protein resulted in selectively increased Il6
expression, as opposed to Il1b and Tnfa transcription. Collectively,
the authors concluded that a combination of Fas up-regulation and IL-6
production by NP+ CD169+ macrophages induce AICD in lymphocytes in
secondary lymphoid organs, resulting in lymphocytopenia.
In summary, this study reports that CD169+ macrophages in the splenic
MZ, subscapular LN, and the lining capillaries of the secondary lymphoid
tissues express ACE2 and are susceptible to SARS-CoV-2 infection. The
findings point to the potential role of these macrophages in viral
dissemination, immunopathology of these secondary lymphoid organs,
hyperinflammation and lymphopenia.
Limitations
Technical
A notable technical limitation is the small number of samples (n=6);
moreover, the analysis of these samples using multiplexed
immunohistochemistry and immunofluorescence do not necessarily provide
the depth of unbiased interrogation needed to better identify the cell
types involved.
Biological
The available literature and ongoing unpublished studies, including
single-cell experiments of spleen and LN from organ donors, do not
indicate that ACE2 is expressed by macrophages; however, it remains
possible that ACE2 expression may be triggered by type I IFN in COVID-19
patients. Importantly, the SARS-CoV-2 NP staining of the macrophages
does not necessarily reflect direct infection of these macrophages;
instead, positive staining only indicates that these macrophages carry
SARS-CoV-2 NP as antigen cargo, which may have been phagocytosed. Direct
viral culture of macrophages isolated from the secondary lymphoid organs
with SARS-CoV-2 is required to confirm the potential for direct
infection of macrophages by SARS-CoV-2. Additionally, it is important to
note that the low to negligible viremia reported in COVID-19 patients
to-date does not favor a dissemination route via the blood, as suggested
by this study, which would be necessary to explain the presence of
virally infected cells in the spleen.
Relevance
Excess inflammation in response to SARS-CoV-2 infection is characterized
by cytokine storm in many COVID-19 patients. The contribution of this
pathology to the overall fatality rate due to COVID-19, not even
necessarily directly due to SARS-CoV-2 infection, is significant. A
better understanding of the full effect and source of some of these
major cytokines, like IL-6, as well as the deficient immune responses,
like lymphocytopenia, is urgently needed. In this study, the authors
report severe tissue damage in spleens and lymph nodes of COVID-19
patients and identify the role that CD169+ macrophages may play in the
hyperinflammation and lymphocytopenia that are both characteristic of
the disease. It may, therefore, be important to note the effects that
IL-6 inhibitors like Tocilizumab and Sarilumab may specifically have on
splenic and LN function. It is important to note that similar
observations of severe splenic and LN necrosis and inflammation in
patients infected with SARS-CoV-1 further support the potential
importance and relevance of this study.
13.92 Cigarette smoke triggers the expansion of a subpopulation of respiratory epithelial cells that express the SARS-CoV-2 receptor ACE2
Study uses scRNAseq, bulk seq data and air-liquid interface culture
experiments to show that cigarette smoke causes a dose-dependent
upregulation of ACE2 in mouse and human lungs (transplantation,
tumor resection, or IPF datasets).
ACE2 was not up-regulated in patients with asthma or
lung-sarcoidosis or in mouse models of cystic fibrosis or carcinogen
exposure.
Cathepsin B (alternate protease involved in viral entry) is
increased in smoke-exposed mouse or human lungs.
Smoke triggers a protective expansion of mucus-secreting MUC5AC+
goblet and SCGB1A1+ club cells; ACE2 presence in these cells is
increased upon smoke exposure.
13.92.3 Limitations:
Long-term smokers usually have several co-morbidities including
immune dysfunction, which can affect interpretation of CoV-2
susceptibility in these datasets. Ideally, analyses can control for
major co-morbidities across smokers and non-smokers (immune
suppression, cardiovascular disease and atherosclerosis).
Hyperplasia of ACE2+ goblet cells upon smoking needs to be separated
from ACE2 upregulation in existing goblet cells.
ACE2 expression increase alone does not confirm increased viral
entry into goblet cells; future studies with air-liquid interface
cultures testing CoV-2 infectivity in ex vivo epithelial cells
from human epithelial lines, ex vivo samples or hACE2 mice will be
very informative.
13.92.4 Significance
This study may partially explain why smokers are more likely to
develop severe SARS-CoV-2 infections. Also, the reversibility of
ACE2 expression upon smoking cessation suggests that quitting
smoking could lessen CoV-2 susceptibility.
Absence of ACE2 upregulation in other lung inflammation pathologies
implies CoV-2 susceptibility might be smoking-specific, and not
fibrosis-specific.
Another preprint showed ACE2 expression increases in lung of
patients with CoV-2 co-morbidities such as hypertension [1343]; these studies
collectively paint a better picture of CoV-2 susceptibility before
actual experiments can be carried out.
13.92.5 Credit
Review by Samarth Hegde as part of a project by students, postdocs and faculty at the Immunology Institute of the Icahn School of Medicine, Mount Sinai.
13.93 The comparative superiority of IgM-IgG antibody test to real-time reverse transcriptase PCR detection for SARS-CoV-2 infection diagnosis
The study compares IgM and IgG antibody testing to RT-PCR detection of
SARS-CoV-2 infection. 133 patients diagnosed with SARS-CoV-2 in Renmin
Hospital (Wuhan University, China) were analyzed. The positive ratio was
78.95% (105/133) in IgM antibody test (SARS-CoV-2 antibody detection kit
from YHLO Biotech) and 68.42% (91/133) in RT-PCR (SARS-CoV-2 ORF1ab/N
qPCR detection kit). There were no differences in the sensitivity of
SARS-CO-V2 diagnosis in patients grouped according to disease severity.
For example, IgG responses were detected in 93.18% of moderate cases,
100% of severe cases and 97.3% of critical cases. In sum, positive
ratios were higher in antibody testing compared to RT-PCR detection,
demonstrating a higher detection sensitivity of IgM-IgG testing for
patients hospitalized with COVID-19 symptoms.
13.93.3 Limitations
This analysis only included one-time point of 133 hospitalized patients,
and the time from symptom onset was not described. There was no
discussion about specificity of the tests and no healthy controls were
included. It would be important to perform similar studies with more
patients, including younger age groups and patients with mild symptoms
as well as asymptomatic individuals. It is critical to determine how
early after infection/symptom onset antibodies can be detected and the
duration of this immune response.
13.93.4 Significance
The IgM-IgG combined testing is important to improve clinical
sensitivity and diagnose COVID-19 patients. The combined antibody test
shows higher sensitivity than individual IgM and IgG tests or nucleic
acid-based methods, at least in patients hospitalized with symptoms.
13.93.5 Credit
Review by Erica Dalla as part of a project by students, postdocs and
faculty at the Immunology Institute of the Icahn School of Medicine,
Mount Sinai.
Title: Lectin-like Intestinal Defensin Inhibits 2019-nCoV Spike
binding to ACE2
Human ACE2 was previously identified as the host receptor for
SARS-CoV-2. Despite ACE2 being expressed in both lung alveolar
epithelial cells and small intestine enterocytes, respiratory problems
are the most common symptom after viral infection while intestinal
symptoms are much less frequent. Thus, the authors here investigate the
biology behind the observed protection of the intestinal epithelium from
SARS-CoV-2. Human defensin 5 (HD5), produced by Paneth cells in the
small intestine, was shown to interact with human ACE2, with a binding
affinity of 39.3 nM by biolayer interferometry (BLI). A blocking
experiment using different doses of HD5 coating ACE2 showed that HD5
lowered viral spike protein S1 binding to ACE2. Further, a molecular
dynamic simulation demonstrated a strong intermolecular interaction
between HD5 and the ACE2 ligand binding domain. To test HD5 inhibitory
effect on S1 binding to ACE2, human intestinal epithelium Caco-2 cells
were preincubated with HD5. Preincubation strongly reduced adherence of
S1 to surface of cells. HD5 was effective at a concentration as low as
10 µg/mL, comparable to the concentration found in the intestinal fluid.
Limitations:
The study focuses exclusively on intestinal cells. However, HD5 could
have been tested to block ACE2-S1 binding in human lung epithelial cells
as a potential treatment strategy. It would be useful to know whether
HD5 could also prevent viral entry in lung cells.
Relevance:
This work provides the first understanding of the different efficiency
of viral entry and infection among ACE2-expressing cells and tissues.
Specifically, the authors show that human defensin 5 produced in the
small intestine is able to block binding between S1 and ACE2 necessary
for viral entry into cells. The study provides a plausible explanation
on why few patients show intestinal symptoms and suggests that patients
with intestinal disease that decrease defensins’ production may be more
susceptible to SARS-CoV-2. It also indicates that HD5 could be used as a
molecule to be exogenously administered to patients to prevent viral
infection in lung epithelial cells.
Title:
Susceptibility of ferrets, cats, dogs and different domestic animals
to SARS-coronavirus-2
This study evaluated the susceptibility of different model laboratory
animals (ferrets), as well as companion (cats and dogs), and domestic
animals (pigs, chickens and ducks) to SARS-CoV-2. They tested infection
with two SARS-CoV2 isolates, one from an environmental sample collected
in the Huanan Seafood Market in Wuhan (F13-E) and the other from a human
patient in Wuhan (CTan-H).
Ferrets were inoculated with either of the two viruses by intranasal
route with 105 pfu, and the viral replication was evaluated. Two
ferrets from each group were euthanized on day 4 post infection (p.i.).
AT day 4 p.i., viral RNA and infectious viruses were detected only in
upper respiratory tract (nasal turbinate, upper palate, tonisls, but not
in the trachea, lungs or other tissues. Viral RNA and virus titer in the
remaining ferrets were monitored in nasal washes and rectal swabs on
days 2, 4, 6, 8 and 10 p.i. Viral RNA and infectious viruses were
detected in nasal washes until day 8 p.i. One ferret in each group
developed fever and loss of appetite on days 10 and 12 p.i., however,
viral RNA was practically undetactable. These two ferrets showed severe
lymphoplasmacytic perivasculitis and vasculitis in the lungs and lower
antibody titers compare to other 4 ferrets.
Cats. Five subadult 8-month-old domestic cats were inoculated with
CTan-h virus and three uninfected cats were placed in a cage adjacent to
each of the infected cats to monitor respiratory droplet transmission.
Viral RNA was detected in the upper respiratory organs from all infected
cats and in one out of three exposed cats. All infected (inoculated and
exposed) cats developed elevated antibodies against SARS-CoV2. Viral
replication studies with juvenile cats (70-100 days) revealed massive
lesions in the nasal and tracheal mucosa epithelium and lungs of two
inoculated cats which died or were euthanized on day 3 p.i., and
infection in one out of three exposed cats. These results indicated
SARS-CoV2 could replicate in cats, that juvenile cats were more
susceptible that adults, and theat SARS-CoV2 could be transmit via
respiratory droplets between cats.
Dogs and others. Five 3-month-old beagle dogs were inoculated and
housed with two uninoculated beagles in a room. Two virus inoculated
dogs seroconverted, but others including two contact dogs were all
seronegative for SARS-CoV2 and infectious virus was not detected in any
swabs collected. Viral RNA was not detected in swabs from pigs,
chickens, and ducks inoculated or contacted. These results indicated
that dogs, pigs, chickens, and ducks might have low or no susceptibility
to SARS-CoV2.
Critical analysis of the study:
This manuscript describes the viral replication and clinical symptoms of
SARS-CoV2 infection in ferrets, and the SARS-CoV2 infection and
transmission in cats. Clinical and pathological analysis was not
performed in cats, therefore the correlation of virus titer with
symptoms severity in the adult and juvenile cats could not be
determined.
The importance and implications for the current epidemics:
SARS-CoV-2 transmission to tigers, cats and dogs has been previously
reported. It should be noted that this manuscript did not evaluate the
transmission from cats to human. Nevertheless, it clearly showed higher
susceptibility of ferrets and domestic cats to SARS-CoV-2. This data
strongly indicates the need for surveillance of possible infection and
transmission of SARS-CoV-2 by domestic cats.
13.94 Virus-host interactome and proteomic survey of PMBCs from COVID-19 patients reveal potential virulence factors influencing SARS-CoV-2 pathogenesis
The authors identified intra-viral protein-protein interactions
(PPI) with two different approaches: genome wide yeast-two hybrid (Y2H)
and co-immunoprecipitation (co-IP). A total of 58 distinct PPI were
characterized. A screen of viral-host PPI was also established by
overexpressing all the SARS-CoV-2 genes with a Flag epitope into HEK293
cells and purifying each protein complex. Interacting host proteins were
then identified by liquid chromatography and tandem mass spectrometry.
251 cellular proteins were identified, such as subunits of ATPase, 40S
ribosomal proteins, T complex proteins and proteasome related proteins,
for a total of 631 viral-host PPI. Several interactions suggesting
protein-mediated modulation of the immune response were identified,
highlighting the multiple ways SARS-CoV-2 might reprogram infected
cells.
Subsequently, the authors compared global proteome profiles of PBMCs
from healthy donors (n=6) with PBMC from COVID-19 patients with mild
(n=22) or severe (n=13) symptoms. 220 proteins were found to be
differentially expressed between healthy donors and mild COVID-19
patients, and a pathway analysis showed a general activation of the
innate immune response. 553 proteins were differentially expressed
between the PBMC of mild and severe COVID-19 patients, most of them
(95%) being downregulated in severe patients. Functional pathway
analysis indicated a defect of T cell activation and function in severe
COVID-19. There was also evidence suggesting reduced antibody secretion
by B cells. Together, these results suggest a functional decline of
adaptive immunity. A FACS analysis of PBMC from severe patients
indicated higher levels of IL6 and IL8 but not IL17 compared to mild
patients.
Finally, the authors focused on NKRF, an endogenous repressor of IL8/IL6
synthesis that was previously identified as interacting with SARS-Cov-2
nsp9,10,12,13 and 15. Individually expressed nsp9 and nsp10 (but not
nsp12, nsp13, nsp15) induced both IL6 and IL8 in lung epithelial A459
cells, indicating that nsp9 and nsp10 may be directly involved in the
induction of these pro-inflammatory cytokines. The authors finally argue
that nsp9 and nsp10 represent potential drug targets to prevent
over-production of IL6 and IL8 in infected cells, and reducing the
over-activation of neutrophils.
13.94.3 Limitations
First, the authors seem to have forgotten to include the extended data
in the manuscript, and their proteomic data does not seem to be publicly
available for the moment, which limits greatly our analysis of their
results.
While this work provides important data on host and viral PPI, only 19
interactions were identified by Y2H system but 52 with co-IP. The
authors do not comment about what could lead to such differences between
the two techniques and they don’t specify whether they detected the same
interactions using the two techniques.
Moreover, the PBMC protein quantification was performed comparing bulk
PBMC. Consequently, protein differences likely reflect differences in
cell populations rather than cell-intrinsic differences in protein
expression. While this analysis is still interesting, a similar
experiment performed on pre-sorted specific cell populations would allow
measuring proteome dynamics at a higher resolution.
Finally, the authors did not discussed their results in regards to
another SARS-CoV-2 interactome of host-viral PPI that had been published
previously1. This study reported 332 host-virus PPI, but no
interaction of viral proteins with NKRF was found. Some interactions
were found in both studies (eg. N and G3BP1, Orf6 and RAE1). However,
the time point used to lyse the cells were different (40h previously vs
72h here), which could explain some of the differences.
13.94.4 Relevance
The identification of many interactions between intra-viral and
host-virus PPI provides an overview of host protein and pathways that
are modulated by SARS-CoV-2, which can lead to the identification of
potential targets for drug development.
In the model proposed by the authors, nsp9 and nsp10 from SARS-Cov-2
induce an over-expression of IL6 and IL8 by lung epithelial cells, which
recruits neutrophils and could lead to an excess in lung infiltration.
Nsp9 has been shown to be essential for viral replication for
SARS-Cov-12, and shares a 97% homology with nsp9 from SARS-Cov-23.
Further, nsp9 crystal structure was recently solved3, which can help
to develop drug inhibitors if this protein is further confirmed as being
important for the virulence of SARS-Cov-2.
1. Gordon DE, Jang GM, Bouhaddou M, et al. A SARS-CoV-2-Human
Protein-Protein Interaction Map Reveals Drug Targets and Potential
Drug-Repurposing. bioRxiv. March 2020:2020.03.22.002386.
doi:10.1101/2020.03.22.002386
2. Miknis ZJ, Donaldson EF, Umland TC, Rimmer RA, Baric RS, Schultz LW.
Severe acute respiratory syndrome coronavirus nsp9 dimerization is
essential for efficient viral growth. J Virol. 2009;83(7):3007-3018.
doi:10.1128/JVI.01505-08
3. Littler DR, Gully BS, Colson RN, Rossjohn J. Crystal Structure of
the SARS-CoV-2 Non-Structural Protein 9, Nsp9. Molecular Biology; 2020.
doi:10.1101/2020.03.28.013920
Title: Prediction and Evolution of B Cell Epitopes of Surface
Protein in SARS-CoV-2
Lon et al. used a bioinformatic analysis of the published SARS-CoV-2
genomes in order to identify conserved linear and conformational B cell
epitopes found on the spike (S), envelope (E), and membrane (M)
proteins. The characterization of the surface proteins in this study
began with an assessment of the peptide sequences in order to identify
hydrophilicity indices and protein instability indices using the
Port-Param tool in ExPASy. All three surface proteins were calculated to
have an instability score under 40 indicating that they were stable.
Linear epitopes were identified on the basis of surface probability and
antigenicity, excluding regions of glycosylation. Using BepiPred 2.0
(with a cutoff value of 0.35) and ABCpred (with a cutoff value of 0.51),
4 linear B cell epitopes were predicted for the S protein, 1 epitope for
the E protein, and 1 epitope for the M protein. For structural analysis,
SARS-CoV assemblies published in the Protein Data Bank (PDB) acting as
scaffolds for the SARS-CoV-2 S and E amino acid sequences were used for
input into the SWISS-MODEL server in order to generate three-dimensional
structural models for the assessment of conformational epitopes. Using
Ellipro (cutoff value of 0.063) and SEPPA (cutoff value of 0.5), 1
conformational epitope was identified for the S protein and 1 epitope
was identified for the E protein, both of which are accessible on the
surface of the virus. Finally, the Consurf Server was used to assess the
conservation of these epitopes. All epitopes were conserved across the
published SARS-CoV-2 genomes and one epitope of the spike protein was
predicted to be the most stable across coronavirus phylogeny.
Critical Analysis/Limitations:
While this study provides a preliminary identification of potential
linear and conformational B cell epitopes, the translational value of
the epitopes described still needs extensive experimental validation to
ascertain whether these elicit a humoral immune response. The
conformational epitope analyses are also limited by the fact that they
are based off of predicted 3D structure from homology comparisons and
not direct crystal structures of the proteins themselves. Additionally,
since there was not a published M protein with a high homology to
SARS-CoV-2, no conformational epitopes were assessed for this protein.
Finally, while evolutionary conservation is an important consideration
in understanding the biology of the virus, conservation does not
necessarily imply that these sites neutralize the virus or aid in
non-neutralizing in vivo protection.
Relevance/Implications:
With further experimental validation that confirms that these epitopes
induce effective antibody responses to the virus, the epitopes described
can be used for the development of treatments and vaccines as well as
better characterize the viral structure to more deeply understand
pathogenesis.
14 Appendix B
Contributors were asked to complete this template to summarize and evaluate new papers related to diagnostics.
Title: Please edit the title to add the name of the paper after the colon
Please paste a link to the paper or a citation here:
Please list some keywords (3-10) that help identify the relevance of this paper to COVID-19
keyword 1 (replace me, copy and paste more than three if needed)
keyword 2 (replace me, copy and paste more than three if needed)
keyword 3 (replace me, copy and paste more than three if needed)
Please note the publication / review status
Pre-print
New Peer-Reviewed Paper
Peer-Reviewed Paper Pre-2020
Which areas of expertise are particularly relevant to the paper?
virology
epidemiology
biostatistics
immunology
pharmacology
Questions to answer about each paper:
Please provide 1-2 sentences introducing the study and its main findings
Study question(s) being investigated:
What type of testing scenario is being considered?
Is it a screening test (used for individuals with no symptoms), diagnostic test (used for individuals with symptoms), or definitive test (used for individuals who have had previous positive test results on diagnostic or screening tests)?
Study population:
What is the model system (e.g., human study, animal model, cell line study)?
What is the sample size?
What is the “pre-test” probability of disease in the study population (i.e., what is the anticipated prevalence of the disease?)
For human studies, the following are related to the pre-test probability:
What countries/regions are considered?
What is the age range, gender, other relevant characteristics?
What is the setting of the study (e.g., random sample of school children, retirement communities, etc.)?
What other specific inclusion-exclusion criteria are considered?
Reference test:
What reference test is considered as a “gold standard” comparator for the test under investigation?
Test assignment:
How are the new and reference tests assigned?
Examples of assignment could include: Recruited individuals have initially undergone neither the new nor the reference test; individuals tested as positive or negative by the reference test undergo the new test; individuals who have undertaken the new test are assessed by the standard test.
Are there any other relevant details about the study design?
Depending on how individuals are chosen, the test may be biasing towards more sick or less sick individuals or very clear-cut positive/negative cases.
Any factors that would influence this bias should be included here.
Test conduct:
How were tests performed?
Describe technical details of assays used, when measurements were taken and by whom, etc. for both the new and standard tests.
Test Assessment
Describe how individuals are classified as positive or negative, e.g. if a threshold is used.
Is there evidence that the test is precise/reproducible when repeated more than once?
Are measurements complete?
For example: Do some participants undergo just one test (the new or the reference test)?
Are there individuals with inconclusive results?
Results summary:
What are the estimated sensitivity, specificity, positive predictive value (PPV), and negative predicted value (NPV)?
Note that the PPV and NPV represent “post-test” probabilities of disease and are generally more meaningful than sensitivity and specificity.
Sometimes the post-test odds will be given instead.
What are the confidence bounds around these intervals?
Interpretation of results for study population:
How good is the test at ruling in or ruling out a disease based on the post-test probabilities?
Are there identified side affects of the test?
Is patient adherence to the test likely to be an issue?
Extrapolation of conclusions to other groups of individuals
How well is the test likely to work in populations with different pretest odds?
For example, if the prevalence is lower, then the PPV will also be lower, but the NPV will be higher.
How costly is the test?
How difficult is it to perform the test in different settings?
Could the test be combined with other existing tests?
Summary of reliability
1-2 sentences on concluding remarks, including summary of strengths, weaknesses, limitations.
Progress
Check off the components as they are completed. If the component is not applicable, check the box as well.
1-2 sentences introducing the study and its main findings
Describe testing scenario
Describe model system
Sample size
Describe prevalnce of disease
Describe countries/regions are considered
Describe age range, gender, other relevant characteristics
Describe setting of the study
Describe other specific inclusion-exclusion criteria
Describe “gold standard”
Describe how the new and reference tests assigned
Describe other relevant details about the study design
Describe how the tests were performed
Describe how individuals are classified as positive or negative
Describe if test is precise/reproducible
Describe whether measurements are complete
What are the estimated sensitivity, specificity, positive predictive value (PPV), and negative predicted value (NPV)?
What are the confidence bounds around these intervals?
Describe post-test probabilities
Describe side affects of the test
Describe patient adherence
Describe how it will extrapolate
How costly is the test?
How difficult is it to perform the test in different settings?
Could the test be combined with other existing tests?
Summary of reliability
15 Appendix C
Contributors were asked to complete this template to summarize and evaluate new papers related to therapeutics.
Title: Please edit the title to add the name of the paper after the colon
Please paste a link to the paper or a citation here:
Please list some keywords (3-10) that help identify the relevance of this paper to COVID-19
keyword 1 (replace me, copy and paste more than three if needed)
keyword 2 (replace me, copy and paste more than three if needed)
keyword 3 (replace me, copy and paste more than three if needed)
Please note the publication / review status
Pre-print
New Peer-Reviewed Paper
Peer-Reviewed Paper Pre-2020
Which areas of expertise are particularly relevant to the paper?
virology
epidemiology
biostatistics
immunology
pharmacology
Questions to answer about each paper:
Please provide 1-2 sentences introducing the study and its main findings
Study question(s) being investigated:
How many/what drugs/combinations are being considered?
What are the main hypotheses being tested?
Study population:
What is the model system (e.g., human study, animal model, cell line study)?
What is the sample size? If multiple groups are considered, give sample size for each group (including controls).
number treated with treatment A
number treated with treatment B
For human studies:
What countries/regions are considered?
What is the age range, gender, other relevant characteristics?
What is the setting of the study (random sample of school children, inpatient, outpatient, etc)?
What other specific inclusion-exclusion criteria are considered?
For example, do the investigators exclude patients with diagnosed neoplasms or patients over/under a certain age?
Treatment assignment:
How are treatments assigned?
For example, is it an interventional or an observational study?
Is the study randomized?
A study can be interventional but not randomized (e.g., a phase I or II clinical trial is interventional but often not randomized).
Provide other relevant details about the design.
This includes possible treatment stratification (e.g., within litters for animal studies, within hospitals for human studies), possible confounding variables (e.g., having a large age range of individuals), possible risks of bias and how they are addressed (e.g., is there masking in a clinical trial? how are individuals chosen in an observational study?).
Outcome Assessment:
Describe the outcome that is assessed and whether it is appropriate.
For example: Is the outcome assessed by a clinician or is it self-reported?
Is the outcome based on viral load or a functional measurement (e.g., respiratory function, discharge from hospital)?
What method is used to measure the outcome?
How long after a treatment is the outcome measured?
Are outcome measurements complete?
For example, are there individuals lost to follow up?
Are outcome measurements subject to various kinds of bias?
For example, a lack of masking in randomized clinical trials.
Statistical Methods Assessment:
What methods are used for inference?
For example, logistic regression, nonparametric methods.
Are the methods appropriate for the study?
For example, are clustered data treated independently or are clusters adjusted for, such as different hospitals or litters?
Are adjustments made for possible confounders?
For example, adjustment for age, sex, or comorbidities.
Results Summary:
What is the estimated association?
For example, is it an estimated odds ratio, a median difference in detected cases, etc?
What measures of confidence or statistical significance are provided?
For example, confidence intervals, p-values, and/or Bayes factors.
Interpretation of results for study population:
Can we make a causal interpretation for the individuals in the study of drug -> outcome, such as “taking drug A improves likelihood of survival twofold over taking drug B.”
For example, with a well-performed animal study or randomized trial it is often possible to infer causality.
If is an observational study, does it match up with some of the Bradford Hill criteria? https://www.edwardtufte.com/tufte/hill https://en.wikipedia.org/wiki/Bradford_Hill_criteria
Are there identified side effects or interactions with other drugs?
For example, is the treatment known to cause liver damage or to not be prescribed for individuals with certain comorbities?
Are there specific subgroups with different findings?
For example, do individuals with a specific baseline seem to do particularly well? Be particularly cautious with respect to multiple testing here.
Extrapolation of conclusions to other groups of individuals not specifically included in the study:
If the study is an animal study, which animal and how relevant is that model?
Is the model system appropriate? Is there evidence from past use that it’s highly-relevant to therapeutic design in this context?
If it is a human study, what characteristics of the study population may support/limit extrapolation?
Can results extrapolate easily to other similar groups? (e.g., same country, similar age groups)
What would happen if conditions are extended in terms of dose or duration?
Can results be extrapolated to other populations or in very different settings? (e.g., different age group, primary care setting vs emergency department etc)
Summary of reliability
1-2 sentences on concluding remarks, including summary of strengths, weaknesses, limitations.
Progress
Check off the components as they are completed. If the component is not applicable, check the box as well.
1-2 sentences introducing the study and its main findings
Describe How many/what drugs/combinations are being considered
Describe the model system
What is the sample size?
What countries/regions are considered
What is the age range, gender, other relevant characteristics
Describe study setting
Describe other specific inclusion-exclusion criteria
Describe how treatments are assigned
Describe randomization (or not) and other relavent details about the design
Describe the outcome that is assessed and whether it is appropriate.
Describe whether the outcome measurements are complete
Are outcome measurements subject to various kinds of bias?
Describe methods used for inference
Describe whether the methods are appropriate for the study
Are adjustments made for possible confounders?
Describe the estimated association
What measures of confidence or statistical significance are provided?
Describe whether a causal interpretation can be made
Are there identified side effects or interactions with other drugs?
Are there specific subgroups with different findings?
If the study is an animal study, which animal and how relevant is that model?
If it is a human study, what characteristics of the study population may support/limit extrapolation?
Summary of reliability
16 Appendix D
Contributors were asked to complete this template to summarize and evaluate new papers related to topics besides therapeutics and diagnostics.
Title: Please edit the title to add the name of the paper after the colon.
General Information
Please paste a link to the paper or a citation here:
Is this paper primarily relevant to Background or Pathogenesis?
Background
Pathogenesis
Methods
Please list some keywords (3-10) that help identify the relevance of this paper to COVID-19
keyword 1 (replace me, copy and paste more than three if needed)
keyword 2 (replace me, copy and paste more than three if needed)
keyword 3 (replace me, copy and paste more than three if needed)
Please note the publication / review status
Pre-print
New Peer-Reviewed Paper
Peer-Reviewed Paper Pre-2020
Which areas of expertise are particularly relevant to the paper?
virology
epidemiology
biostatistics
immunology
pharmacology
other:
Summary
Suggested questions to answer about each paper:
- What did they analyze?
- What methods did they use?
- Does this paper study COVID-19, SARS-CoV-2, or a related disease and/or virus?
- What is the main finding (or a few main takeaways)?
- What does this paper tell us about the background and/or diagnostics/therapeutics for COVID-19 / SARS-CoV-2?
- Do you have any concerns about methodology or the interpretation of these results beyond this analysis?
 0000-0001-7688-1770 ·
0000-0001-7688-1770 ·  rando2 ·
rando2 ·  tamefoxtime
tamefoxtime